ОЦЕНКА НАСЛЕДОВАНИЯ КОЛИЧЕСТВЕННЫХ ПРИЗНАКОВ
1) Методы оценки количественных признаков и расчет коэффициента наследуемости в широком смысле H2
Лесным селекционерам очень важно знать, наследуются ли признаки, с которыми ему предстоит работать, или нет. При этом чаще всего лесоводы работают с признаками, которые можно измерить: высота и диаметры, приросты, масса плодов и семян, величина их, масляничность и содержание белков у орехоплодовых, длина волокна у ели, пихты, осины. Признаки, которые можно измерить, называются количественными ( в отличие от качественных – окраска, цвет, опушенность и т. д.). Количественные признаки контролируются чаще всего неаллельными, аддитивными (суммарными) генами, и они контролируют практически все хозяйственно ценные признаки. Для того чтобы определить, наследуется ли данный признак или нет, пользуются методами математической статистики, в основе которой лежит теория вероятностей. При этом чаще всего пользуются методом дисперсионного анализа, определяя долю наследственных (генотипических) и ненаследственных (паратипических, экологических) факторов в изменчивости признака. Исследуемые признаки проявляются через фенотипы, поэтому можно сказать, что фенотипическая дисперсия S2ph равна сумме дисперсий генотипической S2g и экологической S2e:
S2ph= S2g+ S2e
Дисперсия рассчитывается по формуле
S2 = 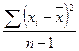 ,
,
где S2 – варианса для выборки,
 – среднее значение признака,
– среднее значение признака,
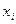 – любой член вариационного ряда,
– любой член вариационного ряда,
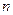 – объем выборочной совокупности выборки (или число измерений).
– объем выборочной совокупности выборки (или число измерений).
Наиболее просто наследуемость того или иного количественного признака может быть определена в популяции, состоящий из нескольких различающихся в генотипическом отношении клонов. На одном участке выращивается несколько клонов (черенкованных или привитых), а затем сравнивают изменчивость между клонами и внутри них. Вследствие того, что внутри отдельного клона генотипическая изменчивость отсутствует, то S2g= 0, исходя из формулы (1) будем иметь S2ph= S2e, то есть внутри клона фенотипическая дисперсия признака равна экологической. Внутри популяции, то есть между клонами, фенотипическая варианса определяется по всем растениям. Средняя же изменчивость признака внутри клонов дает экологическую дисперсию. Тогда доля генотипической дисперсии в общей фенотипической равна
S2ph - S2e .
S2ph
Она обозначается через H2 (heritability – наследуемость) и называется коэффициентом наследуемости в широком смысле, то есть рассчитав ее, мы узнаем, наследуется ли данный признак или его изменения зависят от экологических условий .
Задача. Определить коэффициент наследуемости высоты растений в экспериментальной популяции, состоящей из трех клонов тополей (по Петрову,1973) (табл.14).
Таблица 14
Расчет экологической дисперсии
| Клон 1 | Клон 2 | Клон 3 | ||||||||||||||||||||
| Высота растений | 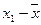
| 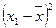
| Высота растений |
|
| Высота растений | 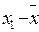
| 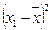
| ||||||||||||||
| –1 | –1 | |||||||||||||||||||||
| –1 | ||||||||||||||||||||||
| –2 | ||||||||||||||||||||||
| –1 | ||||||||||||||||||||||
| –1 | –1 | |||||||||||||||||||||
å = 8 å = 4 å = 8
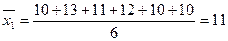 ,
, 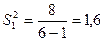 ;
;
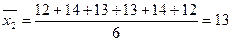 ,
, 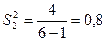 ;
;
 ,
,  ;
;
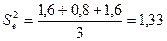 .
.
Таблица 15
Расчет фенотипической дисперсии
| Высота растений в популяциях | 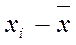
| 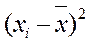
|
| -2 -2 -2 -1 -1 -2 |
å 216 å 32
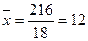 , или
, или 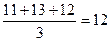 .
.
S2ph= 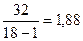 ; H2 =
; H2 = 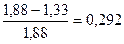 .
.
Вычисляемое значение коэффициента наследуемости свидетельствует о том, что изменчивость высоты растений в изучаемой экспериментальной популяции в значительной большей степени (70,8 %) обусловлена условиями внешней среды, чем наследственными различиями клонов, входящих в состав этой популяции (29,2 %).
Считается, что если H2 > 0,5 (50 %), то признак обусловлен генетически.
Роне В. М. (1980) предложила более простой способ расчета H2 для вегетативно размноженных растений. Генетическая составляющая при этом была обозначена как H2 , а экологическая – е2. Если S2ph=1, тогда S2ph= H2+e2, или 1= H2+е2, то H2=1–е2. Если е2=0, то H2=1, то есть проявление признака не зависит от экологических условий. При е2 не равно 0, выраженность признака в клоне меняется, и сопоставление интервала изменчивости признака в клоне и популяции можно выразить, так:
Lim cl = Xmax – Xmin; Limp=Xmax–Xmin,
– где max и min – значения признака в пределах клона и популяции.
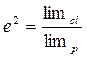 , тогда
, тогда 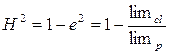 .
.
Этот способ не требует нормального распределения признаков – явления, редкого в природных популяциях.
Рассчитаем предложенную задачу через лимиты:
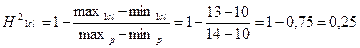 ;
;
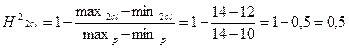 ;
;
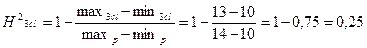 ;
;
 .
.
При расчете коэффициента наследуемости в широком смысле через лимиты H2<0,5, следовательно признак не является генетически обусловленным и в большей степени зависит от экологических факторов.
2) Методика расчета коэффициента наследуемости в узком смысле h2
Узнав, наследуется ли данный признак, селекционер должен знать, какие гены контролируют его. Генотипическая дисперсия S2g представляет собой сумму дисперсий аддитивных генов S2add, эпистатических S2e и доминантных S2d :
S2g = S2add + S2e + S2d.
Райт С. разработал теоретическу модель, выражающую связь фенотипов с генотипами между родственными особями При этом рассчитывается коэффициент наследуемости в узком смысле h2. Он показывает долю аддидивных генов в генотипической составляющей, контролирующих количественные признаки. При этом исходят из следующих допущений:
1) изучаемая популяция находится в состоянии генотипического равновесия;
2) количественные признаки определяются аддидивным действием генов;
3) связь между фенотипом родителей и потомков может быть выражена с помощью коэффициента корреляции, то есть эта связь является прямолинейной.
При этом h2 = 2rxy – для сибсов; h2 = 4rxy – для полусибсов, где r – коэффициент корреляции родитель – потомок; x – потомок; y – родитель. Коэффициент корреляции рассчитывается по формуле
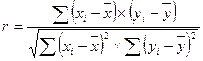 .
.
Задача. Как известно, карельская береза образует несколько габитуальных форм: высокоствольные, короткоствольные, кустарниковые и кустовидные. С каждой из этих форм собраны семена, и выращены в однородных условиях отдельно по семьям сеянцы в количестве 12000 штук. Высоты материнских деревьев у, см и двухлетних сеянцев х, см измерены и средние значения этих показателей представлены в табл. 16.
Таблица 16.
Расчет коэффициента наследуемости в узком смысле h2
| у, см | 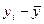
| 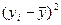
| х, см | 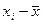
| 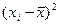
| 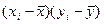
|
| –216,67 | 46945,89 | 11,86 | –3,78 | 14,29 | 819,01 | |
| –166,67 | 27778,89 | 15,74 | 0,1 | 0,01 | –16,67 | |
| 783,33 | 613605,89 | 17,76 | 2,12 | 4,49 | 1660,66 | |
| 183,33 | 33609,89 | 19,24 | 3,6 | 12,96 | 659,99 | |
| –216,67 | 46945,89 | 20,81 | 5,17 | 26,73 | –1120,18 | |
| –316,67 | 100279,89 | 15,61 | –0,03 | 0,0009 | 9,50 | |
| –366,67 | 134446,89 | 13,81 | –1,83 | 3,35 | 671,01 | |
| –166,67 | 27778,89 | 13,26 | –2,38 | 5,66 | 396,67 | |
| 483,33 | 233607,89 | 12,70 | –2,94 | 8,64 | –1420,99 | |
| ∑ 8250 | ∑1265000,01 | ∑140,79 | ∑76,13 | ∑1659 |
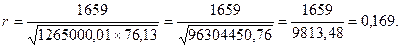
При свободном опылении (полусибсов) полученный коэффициент корреляции умножается на 4.
Таким образом, h2 =4rxy=4×0,169=0,676.
Надежность вычисления коэффициента корреляции зависит от числа саженцев N >100.
Высокий показатель коэффициента наследуемости в узком смысле (h2=0,676) говорит о том, что высоту карельской березы контролируют аддитивные (суммарные) гены, так как h2 > 0,25 (Погиба, 1988). Если h2 < 0,25, то признак контролируется другими генами (Чудный, 1985).