Реакция восстановления.
RCONH2 RCH2NH2; RCONHR' RCH2NHR' ; RCONHR'2 RCH2NHR'2 ;
1. Реакция дегидратации. 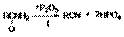
Расщепление по Гофману. 
2. Взаимодействие формамидов с реактивом Гриньяра (реакция Буво)  Продуктами этой реакции являются альдeгид и третичный амин.
Продуктами этой реакции являются альдeгид и третичный амин.
88). Биологическое окисление и окислительное фосфорилирование, тканевое дыхание.Биологическое окисление - это совокупность реакций окисления, протекающих во всех живых клетках. Оно проходит 3 способами: 1.отщепление атомов Н+ 2.присоединение кислорода 3.перенос электронов Тканевое дыхание - это разновидность биологического окисления. В процессе тканевого дыхания в организме вырабатывается АТФ. Тканевое дыхание - это процесс расщепления органических веществ с выделением энергии в виде АТФ, что сопровождается поглощением кислорода, выделением СО2 и образованием воды. Расщепление органических веществ в организме проходит в 3 стадии:
1 и 2 стадия – это специфические пути превращения органических веществ, Ш стадия -–общая, объдиняет все пути превращения органических веществ. В ней образуется наибольшее количество энергии.
Биологические виды энергии. Энергетические превращения в живой клетке подразделяют на две группы: локализованные в мембранах и протекающие в цитоплазме. В каждом случае для «оплаты» энергетических затрат используется своя «валюта»: в мембране это ΔμН+ или ΔμNa+, а вцитоплазме – АТФ, креатинфосфат и другие макроэргические соединения. Непосредственным источником АТФ являются процессы субстратного иокислительного фосфорилирования. Процессы субстратного фосфорилирования наблюдаются при гликолизе и на одной из стадий цикла трикарбоновых кислот (реакция сукцинил-КоА —> сукцинат). Генерация ΔμН+ и ΔμNa , используемых для окислительного фосфорилирования, осуществляется в процессе транспорта электронов в дыхательной цепи энергосопрягающих мембран.Энергия разности потенциалов на сопрягающих мембранах может обратимо превращаться в энергию АТФ. Эти процессы катализируются Н+-АТФ-синтазой в мембранах, генерирующих протонный потенциал, или Na+-АТФ-синтазой (Na+-АТФазой) в «натриевых мембранах» алкалофиль-ных бактерий, поддерживающих ΔμNa+ [Скулачев В.П., 1989]. Cвет или энергиясубстратов дыхания утилизируется ферментами фотосинтетической или дыхательной редокс-цепи (у галобактерий – бактериородопси-ном). Генерируемый потенциал используется для совершения полезной работы, в частности для образования АТФ. Будучи макроэргическим соединением, АТФвыполнняет функцию аккумулирования биологической энергии и ее последующего использования для выполнения клеточных функций. «Макроэргичность» АТФ объясняется рядом особенностей его молекулы.
Тканевое дыхание и биологическое окисление. Распад органических соединений в живых тканях, сопровождающийся потреблением молекулярногокислорода и приводящий к выделению углекислого газа и воды и образованию биологических видов энергии, называется тканевым дыханием. Тканевоедыхание представляют как конечный этап пути превращений моносахаров (в основном глюкозы) до указанных конечных продуктов, в который на разных стадиях включаются другие сахара и их производные, а также промежуточные продукты распада липидов (жирные кислоты), белков (аминокислоты) и нуклеиновых оснований. Итоговая реакция тканевого дыхания будет выглядеть следующим образом: С6Н12О6 + 6O2 = 6СO2+ 6Н2O + 2780 кДж/моль.
В настоящее время биологическое окисление определяется как совокупность реакций окисления субстратов в живых клетках, основная функция которых - энергетическое обеспечениеметаболизма. Потребление кислорода тканями зависит от интенсивности реакций тканевого дыхания. Наибольшей скоростью тканевого дыхания характеризуются почки, мозг, печень, наименьшей – кожа, мышечная ткань (в покое). Уравнение (2) описывает суммарный результат многоступенчатого процесса, приводящего к образованию молочной кислоты и протекающего без участия кислорода: С6Н12Об = 2С3Н6О3 + 65 кДж/моль.
Этот путь отражает, по-видимому, энергетическое обеспечение простейших форм жизни, функционировавших в бескислородных условиях. Современные анаэробные микроорганизмы (осуществляющие молочнокислое, спиртовое и уксуснокислое брожение) получают для жизнедеятельности энергию, производимую в процессе гликолиза или его модификаций.
Использование клетками кислорода открывает возможности для более полного окисления субстратов. В аэробных условиях продукты бескислородного окисления становятся субстратами цикла трикарбоновых кислот , в ходе которого образуются восстановленные дыхательные переносчики НАДФН, НАДН и флавиновые коферменты. Способность НАД+ и НАДФ+ играть роль промежуточного переносчика водородасвязана с наличием в их структуре амида никотиновой кислоты. При взаимодействии этих кофакторов с атомами водорода имеет место обратимое гидрирование (присоединение атомов водорода). При этом в молекулу НАД+ (НАДФ+) включаются 2 электрона и одинпротон, а второй протон остается в среде.
Во флавиновых коферментах (ФАД или ФМН), активной частью молекул которых является изоаллоксазиновое кольцо, в результате восстановлениячаще всего наблюдается присоединение 2 протонов и 2 электронов одновременно.Восстановленные формы этих кофакторов способны транспортироватьводород и электроны к дыхательной цепи митохондрий или иных энергосопрягающих мембран.
Дыхательная цепь - это совокупность ферментов и неферментных компонентов, которые принимают участие в окислении НАДН2 и ФАДН2 и переносе протонов и электронов на кислород.
В состав дыхательной цепи входит НАДН –дегидрогеназа, флавинсодержащие дегидрогеназы, убихинон, цитохромы (цитохромоксидаза), железосерусодержащие белки.
НАДН – ДГ берет участие в окислении НАДН2 .
Убихинон – липофильное вещество, которое свободно плавает в мембране (коэнзим Q). Транспорт электронов и протонов проходит путем передачи двух протонов и двух электронов. Цитохромы находятся в митохондриях. В составе дыхательной цепи можна выделить 4 полиферментных комплекса. В транспорте берут участие железосерусодержащие белки (Fе S Б).
1 система - НАДН –убихиноноксидоредуктаза
П система – сукцинатубихиноноксидоредуктаза
Ш система – убихинолцитохром-С-оксидоредуктаза
1У система – цитохром-С: кислородоксидоредуктаза
В начале цепи энергия протонов и электронов самая большая, а потом по мере транспорта их через различные вещества, она разсеивается и в конце цепи энергия маленькая.
В норме процесс дыхания всегда сопряжен с процессом фосфорилирования, т.е. процессом образования АТФ.
Есть вещества, которые могут угнетать процесс дыхания, они называются ингибиторами. По каждой оксидоредуктазе есть свои ингибиторы. Ими могут быть: ротенон, антимицин, цианиды, чадный газ.
Есть вещества, которые разъединяют процессы окисления и фосфорилирования, они называются разъединителями этих процессов. Ними могут быть физические факторы (холод), химические вещества. Используя ингибиторы, установлено, что в дыхательной цепи есть 3 точки фосфорилирования. Если субстрат окисления НАДН2 , то выделяется 3 молекулы АТФ, если субстрат сукцинат – то выделяется 2 молекулы АТФ. Точка фосфорилирования – это часть дыхательной цепи, в которой выделяется достаточно энергии, которая может быть использована для энергии АТФ.
Сопряжения дыхания и фосфорилирования осуществляется пространственно и энергетически. Пространственно – это все ферменты дыхания и фосфорилирования находятся во внутренней мембране митохондрий. Через внутреннюю: мембрану с помощью транслоказ в матрикс переносятся вещества, которые служат субстратом окисления для дыхательной цепи. Окисление осуществляется только тогда, когда происходит фосфорилирование. Этот процесс контролируется количеством АДФ и АТФ.
В природе есть 2 способа фосфорилирования:
1) субстратное
2) окислительное
Субстратное имеет место 2 раза в гликолизе и один раз в ЦТК. При нем субстрат окисляется, отщепляется е и протоны и на их месте возникает макроэргическая связь.
ГЛИКОЛИЗ – сложный ферментативный процесс последовательных превращений глюкозы, протекающий в тканях человека и животных без потребления кислорода. Конечным продуктом гликолиза является молочная кислота. В процессе гликолиза образуется АТФ.
Процесс может начинаться и с гликогена, тогда он называется гликогенолизом. Другие гексозы (галактоза, фруктоза, сахароза) сначала превращаются в глюкозу, которая затем подвергается гликолизу.
Итак, гликолиз – не только главный путь утилизации глюкозы в клетках, но и уникальный путь, поскольку он может использовать кислород, если последний доступен (аэробные условия), но может протекать и в отсутствие кислорода (анаэробные условия).
Анаэробный распад включает те же реакции специфического пути распада глюкозы до пирувата, но с последующим превращением пирувата в лактат (то есть термины анаэробный распад и анаэробный гликолиз совпадают). Последовательность реакций гликолиза приведена на рисунке:
В гликолизе можно выделить три основных этапа. На первом этапе превращениям подвергаются гексозы, на втором - триозы, на третьем - карбоновые кислоты. Характеристика гликолиза: Характеристика гликолиза:большинство реакций обратимо, за исключением трех (реакций 1, 3, 10); все метаболиты находятся в фосфорилированной форме; источником фосфатной группы в реакциях фосфорилирования являются АТР (реакции 1, 3) или неорганический фосфат (реакция 6); регенерация NAD+, являющаяся необходимым условием протекания гликолиза, происходит при аэробном гликолизе посредством дыхательной цепи. В этом случае водород транспортируется в митохондрии с помощью челночного механизма при участии переносчиков. Это происходит потому, что мембрана митоходрий непроницаема для протонов. При анаэробном гликолизе регенерации NAD+ осуществляется независимо от дыхательной цепи. В этом случае акцептором водорода от NADH является пируват, который восстанавливается в лактат; образование АТР при гликолизе может идти двумя путями: либо субстратным фосфорилированием, когда для фосфорилирования ADP используется энергия макроэргической связи субстрата (реакции 7, 9), либо путем окислительного фосфорилирования ADP, сопряженного с дыхательной цепью (реакция 6).