Активаторы плазминогена выделены из тканей и биологических жидкостей
Активаторы плазминогена (АП)— высокоспецифичные сериновые протеазы регуляторного типа. Известно много АП, выделенных из крови и других биологических жидкостей и тканей человека. Они разделяются на физиологические активаторы, которые в зависимости от источника получения могут быть тканевыми (органными), сосудистыми (тканевый активатор плазминогена), плазменными, кровяными, мочевыми (урокиназа) и т.д. и выделяемые из микроорганизмов (стрептокиназа). Практически все АП образуются в виде проферментов (проактиваторов плазминогена).
Активирование плазминогена может быть:
внешним – под действием активаторов тканей, крови, сосудистой стенки, которые высвобождаются в кровь под влиянием различных факторов;
внутренним – при участии белков плазмы – фактора Хагемана, прекалликреина, высокомолекулярного кининогена;
экзогенным – после введения в организм активаторов плазминогена (стрептокиназа и созданные на ее основе препараты, урокиназа, комплекс стрептокиназа – лиз-плазминоген; тканевый активатор плазминогена, получаемый методом генной инженерии, и другие препараты) с терапевтической целью.
Внутренний путь активации фибринолиза (Хагеманзависимый фибринолиз) инициируется фактором Хагемана (фактор ХП) плазмы крови. После фиксации фактора XII и комплекса высокомолекулярный кининоген-прекалликреин на чужеродной или измененной поверхности (коллаген или др.), путем ограниченного протеолиза происходит образование активного калликреина, который катализирует превращение фактора XII в его активную форму –фактор XIIa. Последний способствует превращению плазминогена в плазмин. Свободный калликреин также является прямым активатором плазминогена.
Хагеманзависимый фибринолиз активируется одновременно с включением каскада реакций образования протромбиназы по внутреннему механизму и его основное назначение – очищение сосудистого русла от фибриновых сгустков, образующихся в процессе внутрисосудистого свертывания крови. В активировании Хагеманзависимого фибринолиза могут принимать участие АПГ, содержащиеся в форменных элементах крови.
Внешний путь активации плазминогена – ведущий путь при повреждении тканей, стимулируется различными тканевыми активаторами плазминогена. Важнейший из них – тканевый активатор плазминогена (тАП), которыйсинтезируется эндотелиальными клетками кровеносных сосудов и по мере надобности расходуется на активацию фибринолиза (рис.13.15).
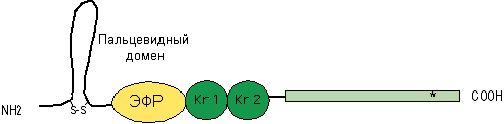
Рис.13.15.Схема строения тАП
Его мол. масса 70 кДа, имеет один домен, структурно подобный ЭФР, 2 крингла и пальцевидный домен, что напоминает структуру плазмина. Секреция тАП эндотелиоцитами происходит не только при тромбозе сосудов, но и при сжатии манжеткой, при физических нагрузках, под влиянием вазоактивных веществ (адреналина, норадреналина) и некоторых лекарственных препаратов. Этот активатор и его ингибиторы обеспечивают постоянно действующую регуляцию фибринолитической активности. На долю тАП приходится 85 % внешней фибринолитической активности крови.
По структуре и механизму действия к тАП близки содержащиеся в разных тканях другие активаторы фибринолиза, которые поступают в кровь при повреждении тканей (травмы, деструкция тканей, акушерская патология и др.). Особое место среди тканевых (органных) факторов фибринолиза занимает продуцируемая почечной тканью и эпителием мочевыводящих путей урокиназа,большая часть которой выделяется с мочой. Урокиназа обеспечивает около 10-15 % внешней фибринолитической активности крови. Она способна проникать внутрь тромба и там катализировать превращение плазминогена в плазмин, разрушая таким образом тромб не только снаружи, но и изнутри.
Кровяные активаторы плазминогенасодержатся в клетках крови (эритроцитах, тромбоцитах и лейкоцитах ) и высвобождаются при их активации и разрушении, а также при тромбообразовании, особенно индуцированном эндотоксином.
Из экзогенных активаторов наиболее изучена стрептокиназа –неферментный белок (мол. м асса 47 кДа), продуцируемый β-гемолитическим стрептококком и в обычных условиях отсутствующий в крови. Стрептокиназа, как и деказа, целиаза, авелизин и другие, не обладают самостоятельной ферментативной активностью по отношению к плазмину, но, соединяясь с плазминогеном, они образуют комплекс, инициирующий превращение плазминогена в плазмин. Таким образом, стрептокиназа активирует плазминоген, связанный с фибриновым сгустком, также, как и плазминоген в растворимой фазе, что сопровождается образованием свободного плазмина. При стрептококковой инфекции возможно образование стрептокиназы в большом количестве, что может приводить к усиленному фибринолизу (фибриногенолизу) и развитию геморрагического диатеза. Превращение плазминогена в плазмин, так же, как и сам процесс лизиса фибриновых сгустков, происходит на поверхности данных сгустков. Сгустки фибрина избирательно адсорбируют и удерживают плазминоген. Богатые лизинами участки (ЛУ), расположенные в центральной части молекулы фибрин(оген)а, связываются с доменами- кринглами плазминогена, при этом одна молекула плазминогена связывается с несколькими молекулами фибрин(оген)а., что позволяет молекуле плазмина действовать на новые интактные молекулы фибрина, оставаясь связанной с субстратом и избегая при этом перехода в раствор и инактивации при контакте с a2-антиплазмином. Вместе с плазминогеном сгусток фибрина специфически связывает активаторы плазминогена. Тканевые активаторы плазминогена имеют низкую каталитическую активность в отсутствие фибрина и активируются при связывании с ним. Активаторы тканевого типа, за исключением урокиназы, имеют более высокое сродство к фибрину в сравнении с фибриногеном, что объясняет преимущественный фибринолиз и в очень слабой степени фибриногенолиз. Одновременное присутствие плазминогена и его активаторов на поверхности фибрина обеспечивает естественное формирование плазмина, и фибрин расщепляется на растворимые фрагменты, получившие название продукты деградации фибрина (ПДФ).
Различные ПДФ проявляют антикоагулянтные, антиполимеризационные, антиагрегационные и другие свойства. Определение ранних и поздних ПДФ проводят для ранней диагностики изменений фибринолитической активности, стадий ДВС-синдромов, дифференциации первичного и вторичного фибринолиза. Ни плазмин, ни активатор плазминогена не связываются с ПДФ и по мере растворения сгустка выходят в плазму, где инактивируются естественными ингибиторами.