Особенности воздействия на клетки-мишени гидрофильных и гидрофобных сигнальных молекул.
Реакция клетки на внеклеточный сигнал осуществляется клетками-мишенями с помощью белков-рецепторов, которые связывают сигнальную молекулу и стимулируют ответную реакцию. Сигнальные молекулы, в зависимости от своей растворимости, связываются с поверхностными или внутриклеточными рецепторами. Жирорастворимые молекулы- стероидные гормоны проходят через плазматическую мембрану и активируют рецепторный белок внутри клетки. Водорастворимые молекулы- большинство гормонов и нейромедиаторы, активируют рецептор на поверхности клеток.
Все водорастворимые и некоторые жирорастворимые сигнальные молекулы присоединяются к специфическим белковым рецепторам на поверхности клеток-мишеней. В зависимости от способа передачи сигнала рецепторы делятся на три класса:
Каналообразующие рецепторы-регулируемые нейромедиаторами ионные каналы, участвующие в быстрой синаптической передаче импульса.
Каталитические рецепторы - при активации лигандом начинают работать как ферменты, фосфорилируя определенные белки. К этой I руппе относятся рецепторы инсулина и многих ростовых факторов.
Рецепторы, сопряженные с G-белками, кот наз вторичными посредниками. Наиболее важные вторичные посредники-циклический АМФ ( ц-АМФ ) и Са2+, кот могут влиять на поведение белков-мишеней, активируя обмен вещ.
Способы передачи сигнала в клетки-мишени от сигнальных молекул:
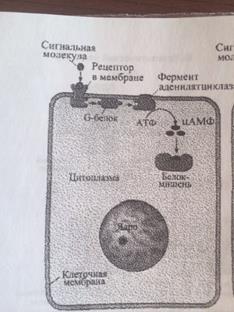
1. АДЕНИЛАТЦИКЛАЗНАЯ (ИЛИ ГУАНИЛАТЦИКЛАЗНАЯ) СИСТЕМА (в мембр скел мышц, жир кл и др)
Сигнальными молекулами для этой системы служат многие гормоны и локальные химические медиаторы. Когда соответствующий лиганд (например, адреналин) взаимодействует с рецептором, происходяi конформационные изменения рецепторного белка. Это вызывао структурную перестройку G-белка, находящегося в мембране. G-белок в этом состоянии активизирует аденилатциклазу - фермент, также располагающийся в мембране. Активизированная аденилатциклаза катализирует превращение АТФ в ц-АМФ. Ц-АМФ воздействует на ферменты протеинкиназы, которые участвуют во многих метаболических процессах в клетке.
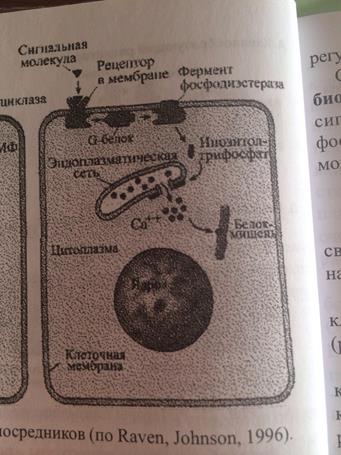
2. ФОСФОИНОЗИТИДНЫЙ МЕХАНИЗМ.
Внутриклеточная система инозитолтрифосфата (ИФ3) имеет сходство с системой ц-АМФ. Как и в системе цАМФ, внеклеточный сигнал опосредован через белок G, который активирует фермент фосфодиэстеразу. При участии этого фермента из фосфолипида внутреннего слоя мембраны образуется инозитолтрифосфат, который диффундирует в цитоплазму. Здесь он вызывает выброс Са2+ из эндоплазматической сети. Повышение концентрации Са2+ в цитоплазме активирует протеинкиназу, которая фосфорилирует и, следовательно, активирует другие ферменты.