ФАКТОРЫ ДИНАМИКИ ЧИСЛЕННОСТИ ПОПУЛЯЦИЙ
Известно три типа зависимости численности популяции от ее плотности (рис. 6). При первом типе (кривая 1) скорость роста популяции уменьшается по мере увеличения плотности. Это широко распространенное явление позволяет понять, почему популяции некоторых животных относительно устойчивы. Во-первых, при увеличении плотности популяции наблюдается снижение рождаемости. Так, в популяции большой синицы при плотности меньше одной пары на 1 га на одно гнездо приходится 14 птенцов; когда же плотность достигает 18 пар на 1 га, выводок составляет менее 8 птенцов. Во-вторых, при увеличении плотности популяции меняется возраст наступления половой зрелости. Например, африканский слон в зависимости от плотности популяции может достигать половой зрелости в возрасте от 12 до 18 лет. Кроме того, этот вид при низкой плотности дает приплод 1 слоненок за 4 года, тогда как при высокой — рождаемость составляет 1 слоненок за 7 лет.
При втором типе зависимости (кривая 2) темп роста популяции максимален при средних, а не при низких значениях плотности. Так, у некоторых видов птиц (например, чаек) число птенцов в выводке увеличивается с повышением плотности популяции, а затем, достигнув наибольшей величины, начинает уменьшаться. Этот тип влияния плотности популяции на скорость размножения особей характерен для видов, у которых отмечен групповой эффект. При третьем типе (кривая 3) темп роста популяции не изменяется до тех пор, пока она не достигнет высокой плотности, затем резко падает.
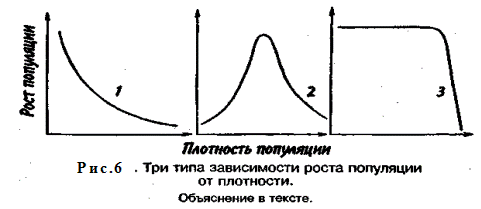
Подобная картина наблюдается, например, у леммингов. При пике численности плотность леммингов становится избыточной, и они начинают мигрировать. Элтон так описал миграции леммингов в Норвегии: животные проходили через деревни в таком количестве, что собаки и кошки, которые вначале нападали на них, просто перестали их замечать. Достигнув моря, обессиленные лемминги тонули.
Регуляция численности равновесных популяций определяется преимущественно биотическими факторами. Среди них главным фактором часто оказывается внутривидовая конкуренция. Примером может служить борьба птиц за мести для гнездования.
Внутривидовая конкуренция может быть причиной физиологического эффекта, известного под названием шоковой болезни. Его отмечают, в частности, у грызунов. Когда плотность популяции становится слишком большой, шоковая болезнь приводит к снижению плодовитости и увеличению смертности, что возвращает плотность популяции к нормальному уровню.
У некоторых видов животных взрослые особи питаются собственным приплодом. Это явление, известное как каннибализм, снижает численность популяции. Каннибализм свойствен, например, окуням: в озерах Западной Сибири 80% пищи крупных особей составляет молодь того же вида. Молодь, в свою очередь, питается планктоном. Таким образом, когда нет других видов рыб, взрослые особи живут за счет планктона.
Межвидовые взаимодействия также играют существенную роль в контроле плотности популяции. Взаимодействия паразит-хозяин и хищник-жертва часто за висят от плотности. Болезни также являются фактором, принимающим участие в регуляции плотности популяции. Когда кролики болеют миксоматозом, вызываемым вирусом, распространение инфекции идет гораздо быстрее в популяциях с повышенной плотностью.
Хищничество как ограничивающий фактор само по себе имеет большое значение. Причем если влияние жертвы на численность популяции хищника не вызывает сомнений, то обратное воздействие, т. е. на популяцию жертвы, бывает не всегда. Во-первых, хищник уничтожает больных животных, тем самым он улучшает средний качественный состав популяции жертвы. Во-вторых, роль хищника ощутима только тогда, когда оба вида обладают приблизительно одинаковым биотическим потенциалом. В противном случае из-за низкого темпа размножения хищник не в состоянии ограничить численность своей жертвы. Например, только одни насекомоядные птицы не могут остановить массовое размножение насекомых. Иными словами, если биотический потенциал хищника намного ниже биотического потенциала жертвы, действие хищника приобретает постоянный характер, не зависящий от плотности его популяции.
Численность насекомых-фитофагов нередко определяется комбинацией видоспецифичных реакций насекомых и растений на воздействие загрязняющих веществ. Загрязнение понижает резистентность растений, в результате чего численность насекомых возрастает. Однако при слишком большом загрязнении численность насекомых падает, несмотря на уменьшение сопротивляемости растений.
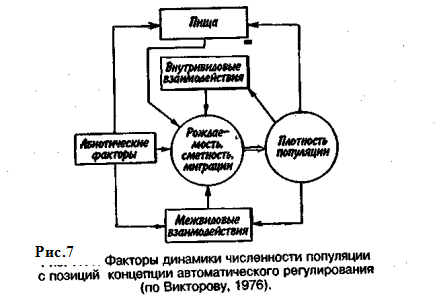
Приведенная дифференциация факторов динамики численности популяций позволяет понять их реальное значение в жизни и воспроизводстве популяций. Современная концепция автоматического регулирования численности популяций базируется на сочетании двух принципиально различных явлений: модификаций, или случайных колебаний численности, ирегуляций, действующих по принципу кибернетической обратной связи и нивелирующих колебания. В соответствии с этим выделяют модифицирующие (независящие от плотности популяции) и регулирующие (зависящие от плотности популяции) экологические факторы, причем первые из них воздействуют на организмы либо непосредственно, либо через изменения других компонентов биоценоза. По существу, модифицирующие факторы представляют собой различные абиотические факторы. Регулирующие факторы связаны с существованием и активностью живых организмов (биотические факторы), поскольку лишь живые существа способны реагировать на плотность своей популяции и популяций других видов по принципу отрицательной обратной связи (рис. 7).
Если воздействия модифицирующих факторов приводят лишь к преобразованиям (модификациям) колебаний численности, не устраняя их, то регулирующие факторы, выравнивая случайные отклонения, стабилизируют (регулируют) численность на определенном уровне. Однако на разных уровнях численности популяции регулирующие факторы принципиально различны (рис. 8). Например, хищники-полифаги, способные при изменении численности жертв ослабить или усилить свою активность—функциональная реакция,—оказывают действие при сравнительно низких значениях численности популяции жертвы.
Хищники - олигофаги, характеризующиеся в отличие от полифагов численной реакцией на состояние популяции жертвы, оказывают на нее регулирующее действие в более широком диапазоне, чем полифаги. При достижении популяцией жертвы еще более высокой численности создаются условия для распространения болезней и, наконец, предельный фактор регуляции — внутривидовая конкуренция, ведущая к исчерпыванию доступных ресурсов и развитию стрессовых реакций в популяции жертвы. На рис. 8 представлена многозвенная буферная система регулирования численности популяции под воздействием биотических факторов, степень влияния которых зависит от плотности популяции. В реальной ситуации данный параметр зависит от большого числа факторов, в частности и тех, которые не оказывают регулирующего воздействия на плотность популяции по принципу обратной связи. Взаимодействие между модифицирующими, регулирующими, а также такими специфическими факторами, как размеры тела, группы и индивидуального участка, при их влиянии на плотность популяции млекопитающих показано на рис. 9.
Таким образом, чтобы получить исчерпывающую информацию о том, какие факторы вызывают колебания численности, теоретически требуются данные о физико-химических условиях, обеспеченности ресурсами, жизненном цикле этих организмов и влиянии конкурентов, хищников, паразитов и т. д., причем нужно знать, как все эти факторы влияют на рождаемость, смертность и миграцию. Все популяции непрерывно изменяются: новые организмы рождаются или прибывают как иммигранты, а прежние гибнут или эмигрируют. Несмотря на это, флуктуации размера популяции не безграничны. С одной стороны, она не может расти беспредельно, а с другой — вымирают виды достаточно редко. Следовательно, один из основных признаков популяционной динамики —сочетание изменений с относительной стабильностью. При этом колебания размеров популяций сильно различаются у разных видов.


Особи в популяции взаимодействуют между собой, обеспечивая свою жизнедеятельность и устойчивое воспроизводство популяции.
У животных, ведущих одиночный образ жизни или создающих семьи, регулирующим фактором является территориальность, которая влияет на обладание определенными пищевыми ресурсами и имеет большое значение для размножения. Особь защищает пространство от вторжения и открывает ее для другой особи только при размножении. Самое рациональное использование пространства достигается в том случае, если образуется настоящая территория — участок, из которого другие особи изгоняются. Так как владелец участка психологически господствует на нем, для изгнания чаще всего достаточно лишь демонстрации угроз, преследования, самое большее — притворных атак, которые прекращаются еще на границах участка. У данных животных огромное значение имеют индивидуальные различия между особями — наиболее приспособленные обладают большим индивидуальным пищевым ареалом.
У животных, ведущих групповой образ жизни и образующих стаи, стада, колонии, групповая защита от врагов и совместная за- бота о потомстве повышает выживаемость особей, что влияет на численность популяции и ее выживаемость. Данные животные организованы иерархически. Иерархические отношения соподчинения построены на том, что ранг каждого известен каждому. Как правило, высший ранг принадлежит самому старшему самцу. Иерархия контролирует все взаимодействия внутри популяции: брачные, особей разных возрастов, родителей и потомства.
У животных особую роль играют отношения «мать—дитя». Родители передают потомкам генетическую информацию и информацию о внешней среде.