Генетический код
Генетический код — способ кодирования аминокислотной последовательности белков при помощи последовательности нуклеотидов. Как только в 1953 г. появилась знаменитая двойная спираль Уотсона и Крика, сразу же возник вопрос: как эта запись прочитывается и используется в клетке? В 1954 году Георгий Гамов сформулировал проблему генетического кода, т.е. задолго до открытия самой мРНК. Размышляя над тем, как линейная последовательность 4-х различных нуклеотидов может определить последовательность 20 разных аминокислот в белке, Гамов предположил, что генетически код является триплетным. Он же поставил вопрос и о других свойствах генетического кода:
1.Триплетность. Одну аминокислоту кодирует последовательность из трех нуклеотидов, названная триплетом, или кодоном.
2. Вырожденность. Каждая аминокислота зашифрована более чем одним кодоном. Исключение составляют аминокислоты метионин и триптофан. Каждая из них кодируется только одним триплетом. Для кодирования 20 аминокислот используется 61 комбинация нуклеотидов. Триплет АУГ, кодирующий метионин, называют стартовым. С него начинается синтез белка. Три кодона (УАА, УАГ, УГА) несут информацию о прекращении синтеза белка. Их называют триплетами терминации.
3. Универсальность. Одни и те же триплеты кодируют одинаковые аминокислоты.
4. Однозначность. Каждый триплет кодирует только одну аминокислоту.
Исключение составляет кодон AUG. У прокариот в первой позиции (заглавная буква) он кодирует формилметионин, а в любой другой - метионин.
5. Колинеарность – совпадение последовательностей аминокислот в синтезируемой молекуле белка с последовательностью триплетов в и–РНК.
6. Непрерывность — между триплетами нет знаков препинания, то есть информация считывается непрерывно.
7. Неперекрываемость — один и тот же нуклеотид не может входить одновременно в состав двух или более триплетов.
8*. Однонаправленность – текст читается только в направлении 5’ 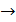 3’.
3’.
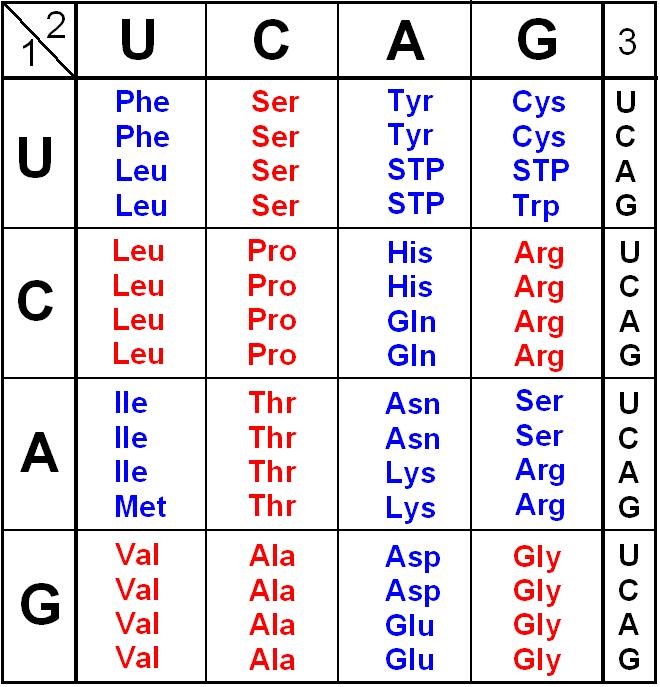
В 1976г. была секвенирована ДНК фага φХ174. У него одноцепочечная кольцевая ДНК, состоящая из 5375 нуклеотидов. Было известно, что фаг кодирует 9 белков. Для 6 из них были определены гены, располагающиеся друг за другом.
Выяснилось, что есть перекрывание. Ген Е полностью находится внутри гена D. Его инициирующий кодон появляется в результате сдвига считывания на один нуклеотид. Ген J начинается там, где кончается ген D. Инициирующий кодон гена J перекрывается с терминирующим кодоном гена D в результате сдвига на два нуклеотида. Конструкция называется "сдвиг рамки считывания" на число нуклеотидов, некратное трем. На сегодняшний день перекрывание показано только для нескольких фагов.
Первый пример отклонения от стандартного генетического кода был открыт в 1979 году при исследовании генов митохондрий человека. С того времени было найдено несколько подобных вариантов, например, прочитывание стоп-кодона УГА в качестве кодона, определяющего триптофан у микоплазм. У бактерий и архей ГУГ и УУГ часто используются как стартовые кодоны. В некоторых случаях гены начинают кодировать белок со старт-кодона, который отличается от обычно используемого данным видом.
Несмотря на эти исключения, у всех живых организмов генетический код имеет общие черты: кодон состоят из трёх нуклеотидов, где два первых являются определяющими, кодоны транслируются тРНК и рибосомами в последовательность аминокислот.
| Кодон | Универсальный код | Митохондриальные коды | |||
| Позвоночные | Беспозвоночные | Дрожжи | Растения | ||
| UGA | STOP | Trp | Trp | Trp | STOP |
| AUA | Ile | Met | Met | Met | Ile |
| CUA | Leu | Leu | Leu | Thr | Leu |
| AGA | Arg | STOP | Ser | Arg | Arg |
| AGG | Arg | STOP | Ser | Arg | Arg |