Тема 2. Методы психофизиологических исследований
Психофизиология относится к экспериментальным дисциплинам, основными методами которой являются электрофизиологические в силу того, что именно физиологические показатели позволяют проникнуть в суть психических процессов и состояний, как на уровне сознания, так и на бессознательном уровне. Электрофизиологические показатели отражают физико-химические следствия обмена веществ, которые сопровождают основные жизненные процессы. Они являются наиболее точными и надежными показателями течения любых физиологических процессов, дают возможность изучения явления или процесса, не травмируя и не искажая его хода и структуры.
В настоящее время в психофизиологических исследованиях все больше делается акцент на изучение нейронных механизмов психических процессов и состояний. Это связано с тем, что все внешние реакции реализуются посредством нейронной активности.
Среди множества электрофизиологических методов, используемых в психофизиологических исследованиях, центральное место занимают различные способы регистрации электрической активности ЦНС и, в первую очередь, головного мозга, безусловно, доминируют такие как: регистрация импульсной активности нервных клеток, ЭЭГ, регистрация вызванных потенциалов мозга человека и потенциалов, связанных с событиями, различные методы томографии, среди которых, прежде всего, следует выделить позитронно-эмиссионную томографию (ПЭТ) и магнитно-резонансную томографию (МРТ). Остановимся на описании некоторых из них.
Электроэнцефалография является одним из основных методов объективного тестирования функций нервной системы.
ЭЭГ - метод исследования головного мозга, основанный на регистрации его электрических потенциалов. Первая публикация о наличие токов в центральной нервной системы была сделана Du Bois Reymond в 1849 г. В 1875 г., данные о наличии спонтанной и вызванной активности в мозге собаки были получены независимо R.Caton в Англии и В.Я.Данилевским в России. Исследования отечественных нейрофизиологов на протяжении конца XIX и начала XX века внесли существенный вклад в разработку электроэнцефалографии. В.Я.Данилевский не только показал возможность регистрации электрической активности мозга, но и подчеркивал ее тесную связь с нейрофизиологическими процессами. В 1912 г. П.Ю.Кауфман выявил связь электрических потенциалов мозга с “внутренней деятельностью мозга” и их зависимость от изменения метаболизма мозга, воздействия внешних раздражений, наркоза и эпилептического припадка. Подробное описание электрических потенциалов мозга собаки с определением их основных параметров было дано в 1913 и 1925 гг. В.В.Правдич-Неминским.
Австрийский психиатр Ганс Бергер в 1928 г. впервые осуществил регистрацию электрических потенциалов головного мозга у человека, используя скальповые игольчатые электроды. В его же работе были описаны основные ритмы ЭЭГ и их изменения при функциональных пробах и патологических изменениях в мозге. Большое влияние на развитие метода оказали публикации G.Walter (1936) о значении ЭЭГ в диагностике опухолей мозга, а также работы F.Gibbs E.Gibbs W.G.Lennox, давших подробную элктроэнцефалографическую семиотику эпилепсии.
В последующие годы работы исследователей были посвящены не только феноменологии электроэнцефалографии при различных заболеваниях и состояниях мозга, но и изучению механизмов генерации электрической активности. Существенный вклад в эту область внесен работами E.D.Adrian, B.Metthew, G.Walter, H.Jasper, В.С.Русинова, В.Е.Майорчик, Н.П.Бехтеревой, Л.А.Новиковой.
В клинической электроэнцефалографии используют две основные системы отведения ЭЭГ: международную систему «10-20» (Jasper H.), а также модифицированные схемы с уменьшенным количеством электродов (Gibbs F., Gibbs E.; Jung J.).
Точки расположения электродов в системе «10-20» определяют следующим образом. Измеряют расстояние по сагиттальной линии от inion до nasion и принимают его за 100%. В 10% этого расстояния от inion и nasion устанавливают соответственно нижний лобный (Fp) и затылочный (О) сагиттальные электроды. Остальные сагиттальные электроды (F, Cz и Р) располагают между этими двумя на равных расстояниях, составляющих 20% от расстояния inion-nasion. Вторая основная линия проходит между двумя слуховыми проходами через vertex (макушку). Нижние височные электроды (ТЗ, Т4) располагают соответственно в 10% этого расстояния над слуховыми проходами, а остальные электроды этой линии (СЗ, Cz, С4) — на равных расстояниях, составляющих 20% длины биаурикулярной линии. Через точки ТЗ, СЗ, С4, Т4 от inion к nasion проводят линии и по ним располагают остальные электроды (РЗ, Р4, Т5, Т6, F3, F4, F7, F8, Fpl, Fp2). На мочки ушей помещают электроды, обозначаемые соответственно А1 и А2. Буквенные символы обозначают основные области мозга и ориентиры на голове: О — occipitalis, Р — parietalis, С — centralis, F — frontalis, A — auricularis. Нечётные цифровые индексы 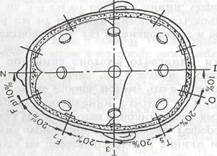 соответствуют электродам над левым, а чётные — над правым полушарием мозга (рис. ).
соответствуют электродам над левым, а чётные — над правым полушарием мозга (рис. ).
| NASION |

Рис.1. Международная схема расположения электродов 10-20
Анализ ЭЭГ основывается на выделении характерных типов электрических потенциалов и определении локализации их источников в мозге. В электроэнцефалографии регистрируется разность потенциалов между двумя точками. С теоретической точки зрения, регистрация потенциала в какой-либо точке в неискаженном виде возможна в условиях, когда один электрод расположен в непосредственной близости от источника потенциала, а другой бесконечно удален от него. В практике используется монополярное и биполярное отведение потенциалов.
Одним из важных описательных терминов является «активность», определяющая любую последовательность волн в ЭЭГ. Характер активности определяется или количественно по частоте и амплитуде (альфа-активность, высокоамплитудная активность и др.) или характеризуется описательно (активность типа спайк-волна, активность типа острых волн).
Для выделения на ЭЭГ значимых признаков её подвергают анализу. Как для всякого колебательного процесса, основными понятиями, на которые опирается характеристика ЭЭГ, являются частота, амплитуда и фаза.
Частота определяется количеством колебаний в секунду, её записывают соответствующим числом и выражают в герцах (Гц). Поскольку ЭЭГ представляет собой вероятностный процесс, на каждом участке записи встречаются, строго говоря, волны различных частот, поэтому приводят среднюю частоту оцениваемой активности. Обычно берут 4-5 отрезков ЭЭГ длительностью 1с и считывают количество волн на каждом из них. Средняя из полученных данных будет характеризовать частоту соответствующей активности на ЭЭГ (рис.1).

|

Рис.2. Измерение частоты (Г) и амплитуды (II) на ЭЭГ.
Частота измеряется как число волн в единицу времени (lc). A — амплитуда.
Амплитуда — размах колебаний электрического потенциала на ЭЭГ, её измеряют от пика предшествующей волны до пика последующей волны в противоположной фазе (см. рис. 13); оценивают амплитуду в микровольтах (мкВ). Для измерения амплитуды используют калибровочный сигнал. Так, если калибровочный сигнал, соответствующий напряжению 50 мкВ, имеет на записи высоту 10 мм, то соответственно 1 мм отклонения пера будет означать 5 мкВ. Измерив циркулем амплитуду волны ЭЭГ в миллиметрах и помножив её на 5 мкВ, получим амплитуду этой волны. В компьютеризированных устройствах значения амплитуд можно получать автоматически. Для характеристики амплитуды активности в описании ЭЭГ принимают наиболее характерно встречающиеся максимальные её значения при монополярном отведении, исключая выскакивающие, которые описывают как особые переходные события.
Фаза определяет текущее состояние процесса и указывает направление вектора его изменений. Некоторые феномены на ЭЭГ оценивают количеством фаз, которые они содержат. Монофазным называется колебание в одном направлении от изоэлектрической линии с возвратом к исходному уровню, двухфазным — такое колебание, когда после завершения одной фазы кривая переходит исходный уровень, отклоняется в противоположном направлении и возвращается к изоэлектрической линии. Полифазными называют колебания, содержащие три и более фаз (рис.2).


Рис.2. Монофазный спайк (1), двухфазное колебание (2), трехфазное (3), полифазное (4).
ЭЭГ представляет собой хаотический процесс, обусловленный суммацией электрических потенциалов, генерируемых многими миллионами нейронов, активность которых, в свою очередь, определяется хаотической синаптической бомбардировкой и, возможно, собственными колебаниями потенциала, обусловленными метаболическими процессами. В связи с этим её спектр оказывается чрезвычайно сложным и широким, хотя количественные соотношения мощностей по каждой из этих частот различны. Достаточно точно спектр мощности ЭЭГ может быть оценён только с применением методов автоматического анализа на электронно-вычислительной машине. Из-за ограниченных перцептивных возможностей человека при визуальном анализе ЭЭГ, применяемом в клинической электроэнцефалографии, целый ряд частот не может быть достаточно точно охарактеризован оператором, так как глаз человека выделяет только некоторые основные частотные полосы, явно присутствующие на ЭЭГ. В соответствии с возможностями ручного анализа введена классификация частот ЭЭГ по некоторым основным диапазонам, которым присвоены названия букв греческого алфавита.
Под понятием "ритм" на ЭЭГ подразумевается определённый тип электрической активности, соответствующий некоторому определенному состоянию мозга и связанный с определёнными церебральными механизмами. Соответственно при описании ритма указывается его частота, типичная для определённого состояния и области мозга, амплитуда и некоторые характерные черты его изменений во времени при изменениях функциональной активности мозга.
Альфа (α) - ритм. Частота 8-13 Гц, амплитуда до 100 мкВ, состоит из волн правильной, почти синусоидальной формы. Регистрируется у 85-95% здоровых взрослых. Лучше всего выражен в затылочных отделах, по направлению кпереди амплитуда его постепенно уменьшается. Наибольшую амплитуду ритм имеет в состоянии спокойного расслабленного бодрствования, особенно при закрытых глазах в затемнённом помещении, а также при медитации и длительной монотонной деятельности. Его амплитуда, хотя и является в среднем относительно постоянным параметром для данного индивидуума, весьма существенно колеблется во времени. Помимо изменений амплитуды, связанных с функциональным состоянием мозга, в большинстве случаев достаточно регулярно наблюдаются спонтанные изменения амплитуды, так называемые модуляции α-ритма, выражающиеся в чередующемся нарастании и снижении амплитуды волн с образованием характерных «веретен», длительность которых чаще всего колеблется от 2 до 8 с.
Если испытуемого отвлечь каким-либо раздражителем, то альфа-ритм десинхронизируется, т.е. заменяется низкоамплитудной высокочастотной ЭЭГ. Этот феномен в литературе обозначается терминами реакция активации, пробуждения или десинхронизации.
При повышении уровня функциональной активности мозга (напряжённое внимание, интенсивная психическая работа, чувство страха, беспокойство) амплитуда α-ритма уменьшается, и часто он полностью исчезает. На ЭЭГ появляется высокочастотная нерегулярная активность. Автоматический анализ показывает, что в спектре мощности ЭЭГ исчезает доминантный пик в области α-ритма и спектр уплощается с равномерным распределением мощности по всем основным частотам, что подтверждает физиологическую интерпретацию этого феномена как десинхронизацию активности нейронов. При кратковременном, внезапно возникающем на фоне покоя внешнем раздражении (особенно вспышке света) эта десинхронизация возникает резко и в случае, если раздражение не носит эмоциогенного характера, наблюдается достаточно быстро (через 0,5-2 с) восстановление α-ритма. Такая реакция на ЭЭГ имеет несколько названий в зависимости от значения, которое в неё вкладывается: «реакция активации», «ориентировочная реакция», «реакция угашения α-ритма», «реакция де-синхронизации». Реакция активации возникает как следствие появления нового фактора в окружающей обстановке, требующего дополнительной мобилизации активности организма и ориентации его в новой ситуации. При повторных предъявлениях одного и того же стимула реакция активации постепенно слабеет и через некоторое время практически полностью угасает. Скорость угасания реакции зависит от субъективной значимости стимула, а также от внутреннего состояния организма, в связи, с чем она может служить в определённых ситуациях мерой оценки состояния мозга. Её наличие или отсутствие может быть дополнительным критерием степени реактивности мозга, показателем влияния нейротропных фармакологических средств.
Бета (β)-ритм. Частота 14-40 Гц, амплитуда до 15 мкВ. Лучше всего β-ритм регистрируется в области передних центральных извилин, однако распространяется и на задние центральные и лобные извилины. На ЭЭГ, отведённой от скальпа, в норме он весьма слабо выражен и в большинстве случаев имеет амплитуду 3-7 мкВ, т.е. всего в 2-3 раза превышает относительный уровень собственных шумов усилителя электроэнцефалографа, а при наличии артефактов ЭМГ может ими полностью маскироваться. β-ритм связан с соматическими, сенсорными и двигательными корковыми механизмами и даёт реакцию на двигательную активацию или тактильную стимуляцию. При выполнении или даже умственном представлении движения β-ритм исчезает в зоне соответствующей корковой проекции.
Активность с частотой 40-70 Гц и амплитудой 5-7 мкВ иногда называют гамма (γ)-ритмом. Международная электроэнцефалографическая классификация рекомендует исключить понятие γ-ритма из употребления и использовать вместо этого выражение «высокочастотный β -ритм».
В последнее время рядом исследований было показано, что при использовании цифрового фильтрования и компьютерного вычисления спектральной мощности в диапазоне 35-45 Гц регистрируется особая, не зависимая от традиционного β-ритма, связанная с познавательными операциями, сенсомоторной интеграцией, распознаванием тактильных стимулов и целенаправленными движениями «активность 40 Гц», оценка топического распределения которой и выраженность при соответствующих пробах дает ценную информацию о нормальных механизмах организации соответствующих высших психических функций.
Мю(μ)-ритм. Частота 8-13 Гц, амплитуда до 50 мкВ. μ-ритм, называемый также wicket (англ.), — аркообразный ритм, имеет параметры, совпадающие с параметрами нормального α-ритма, и отличается от него некоторыми физиологическими свойствами и топографией. Визуально регистрируемый μ-ритм наблюдается у относительно небольшого числа индивидуумов (5-15%) и регистрируется в роландической области, т.е. соответственно распределению μ-ритма. Активируется μ-ритм во время умственной нагрузки и психического напряжения. Связан с тактильным и проприоцептивным раздражением и воображением движения. μ-ритм выражен у слепых, компенсирующих потерю зрения развитием тактильного и двигательного исследования среды.
Аналогично β-ритму, μ-ритм снижается (или в небольшом числе случаев нарастает) по амплитуде при двигательной активации или соматосенсорной стимуляции, в связи с чем его ещё называют «сенсомоторным ритмом». Поскольку он обычно имеет низкую амплитуду и при визуальном анализе в большинстве случаев не отличим от α-ритма.
Помимо этих видов активности в ЭЭГ нормального человека описаны также некоторые другие феномены, в частности, каппа (κ)-ритм и лямбда (λ)-волны. Поскольку они наблюдаются в условиях и при функциональных пробах, обычно не применяемых при стандартном исследовании ЭЭГ, а мозговое происхождение каппа-ритма вообще не доказано, они не имеют диагностического значения.
Выделяют также тета ()- активность с частотой 4-6 Гц и амплитудой более 40 мкВ и дельта (δ)-активность с аналогичной тета-ритму амплитудой и частотой 0,5-3 Гц.
Описанные частотно-амплитудные характеристики ритмов ЭЭГ свойственны здоровому мозгу человека.
Метод вызванных потенциалов. Регистрация вызванных потенциалов (ВП) мозга является объективным и неинвазивным методом тестирования функций ЦНС как человека, так и животных. Внастоящее время метод вызванных потенциалов (ВП) мозга находит все более широкое применение как метод, позволяющий получить объективную информацию о состоянии различных сенсорных систем, например, зрения, слуха, осязания, причем о состоянии не только периферических звеньев, но и центральных. Метод позволяет получить объективную информацию без словесного отчета испытуемого, что особенно важно в случае обследования маленьких детей или больных с различными нарушениями сознания или негативным отношением к обследованию. Метод связан с выделением слабых и сверхслабых изменений электрической активности мозга в ответ на стимул и широко используется, благодаря применению для их регистрации современной электронной техники.
Родоначальниками метода ВП являются английский ученый Кэтон (1875-1877), польский ученый Адольф Бек (1890), русские ученые Ларионов (1897-1899) и Правдич-Неминский (1914, 1929). Сам метод ВП возник в 50 годах, почти на 20 лет позже электроэнцефалографии (ЭЭГ).
Исследование вызванных потенциалов мозга основано на регистрации электрических ответов мозга как на экзогенные события, такие как подача внешнего стимула — зрительного, слухового, чувствительного, так и на эндогенные события, связанные с ожиданием, опознанием, принятием решения и инициацией двигательного ответа.
Для выделения электрических ответов мозга используются приемы, близкие к приемам, применяемым в радиолокации для обнаружения слабых сигналов на фоне шума. Впервые методику когерентного накопления или усреднения для детектирования сигналов мозга применил в 50-х годах английский ученый Dawson. Важнейший прорыв в развитии этого направления произошел в начале 70-х годов, когда американский ученый Jewett показал принципиальную возможность регистрации на поверхности головы ответов от отдаленных структур мозга, прежде всего, слуховых стволовых ядер. Это открытие сразу привлекло внимание аудиологов, а позже и клинических нейрофизиологов, неврологов и нейрохирургов, поскольку оно позволяло неинвазивно получить информацию о функциональном состоянии глубинных стволовых структур мозга. Метод исследования коротколатентных ВП (именно так называется этот вид ВП) получил значительное развитие и стал одним из наиболее информативных методов для оценки функционального состояния и мониторирования различных структур мозга в неврологии, нейрохирургии, педиатрии, отоневрологии.
ВП записываются с электродов, располагаемых на поверхности головы. Применение для записи ВП высокочувствительных усилителей цифровых устройств, позволяет выделить слабые сигналы ЦНС, в 5-100 раз меньшие обычной спонтанной активности головного мозга (ЭЭГ), электромиограммы (ЭМГ) и других биоэлектрических сигналов, за счет усреднения большого числа слабых ответов мозга.
Электрические отклики мозга на зрительные, слуховые или сенсорные стимулы оцениваются по изменению основных параметров ВП — амплитуды и длительности (латентности) различных компонентов ответа, несущих информацию о состоянии тех или иных сенсорных систем.
ВП генерируются теми же нейрональными структурами, что и спонтанная ритмика. Особый вклад в генерацию вносят градуальные потенциалы ВПСП и ТПСП (возбуждающие и тормозные постсинаптические потенциалы) дендритных волокон коры или различных ядер мозга. Суммация синаптических градуальных потенциалов дает основной ответ мозга при регистрации на скальпе. Меньший вклад в регистрацию ВП вносят собственно потенциалы действия. Из широко известных ответов 1 пик стволового акустического ВП является чистым потенциалом действия (ПД) от VIII нерва, генерируемым его дистальной частью. Общий вклад образований мозга, участвующих в генерации ВП, схематично представлен на рис.3.
На примере зрительного анализатора схематично представлены основные элементы ЦНС и типы потенциалов, генерируемых на разном уровне, которые могут вносить вклад в ответ, регистрируемый на поверхности скальпа. ПД — потенциал действия участвует в проведении афферентации. ВПСП и ТПСП — возбуждающий и тормозной постсинаптические дендритные потенциалы, генерируемые в релейных, ассоциативных ядрах мозга и коре.
Периферический афферентный синапсы ядра кора
рецепторный нейрон головного
орган мозга

раздражение ПД синаптические ПД скальповые ВП
потенциалы (ближнего и
(ВПСП, ТПСП) дальнего поля)
генерация ___________ проведение-переключение _____________ проведение-обработка
Рис.3. Механизм генерации ВП. Основные элементы ЦНС, дающие вклад в генерацию ВП, регистрируемых на поверхности скальпа.
Различают вызванные потенциалы ближнего поля («near field») и дальнего поля («far field»). В том случае, когда сигнал ВП отражает информацию от генераторов, находящихся в коре, а электроды располагаются на скальпе в непосредственной близости от этих генераторов, сигнал ВП характеризует ближнее поле, например, в случае волны Р100 зрительного ВП, когда генератором является зрительная кора, а электроды располагаются над затылочной областью. В других случаях, как уже твердо установлено, сигнал ВП может быть от дальнего поля, когда электрод располагается на значительном расстоянии от генераторов сигналов, и сигнал проводится к месту регистрации за счет объемного проведения (аналогично регистрации сигнала ЭКГ электродами, находящимся на значительном расстоянии от сердца). Примером такого механизма генерации ВП являются слуховые ядра ствола мозга, располагающиеся на расстоянии 6-10 см от скальповых электродов, которые регистрируют их активность (Jewett et al., 1970, 1971).
Таким образом, скальповые ВП представляют сумму потенциалов различного типа, в зависимости от отведения и способа регистрации ближнего и дальнего поля.
Основная сложность регистрации ВП заключается в том, что ответы мозга значительно ниже активности спонтанной ритмики ЭЭГ и других сигналов, но имеют с ними общий спектр. Поэтому практически ни один из этих сигналов ВП мы не можем видеть в обычной записи ЭЭГ. Для того чтобы их увидеть и зарегистрировать, нужно, чтобы сигнал ВП превышал спонтанную ритмику и другие шумы, по крайней мере, вдвое.Основным методом, используемым в настоящее время для выделения ВП, является метод синхронного или когерентного накопления (усреднения).
Название этот метод получил из-за того, что при регистрации ВП происходит синхронизация накопления (усреднения) ответов с подаваемым стимулом. Выделение ВП происходит за счет многократной подачи стимулов и суммации каждого последующего ответа с предыдущими. Опорным моментом для такой суммации является момент подачи стимулов. В результате такого накопления сигнал ВП, закономерно связанный со стимулом, растет значительно быстрее, чем шум спонтанной ритмики, не связанный со стимулом и попадающий при суммации в случайную фазу друг с другом. В результате, чем больше число накоплений, тем лучше выделение сигнала из шума.
На рис.4 приведен пример процесса выделения ВП из шума методом синхронного (когерентного) накопления. На верхней кривой 1 показаны отметки стимулов (вспышка света). На кривой 2 сигнал ВП в ЭЭГ при подаче стимула, практически, не виден. На кривой 3 показан ВП в процессе синхронного накопления, по мере увеличения числа суммируемых ответов. Уже после 15 таких накоплений ответ начинает выделяться из спонтанной ритмики, и на нижней кривой показано, что на нем уже могут быть измерены основные параметры ответа.
Обычно, вместо синхронного накопления используют методику синхронного или когерентного усреднения, когда происходит не только сложение ответов, но и деление на число суммации. В большинстве своем в современных устройствах используется именно эта методика, и, поэтому, современные цифровые устройства для выделения ВП называют еще нейроусреднителями.
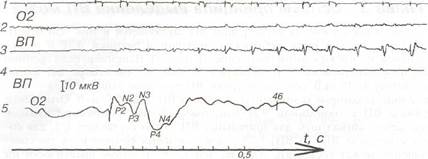
|
Рис.4. Иллюстрация принципа выделения ВП методом синхронного когерентного накопления:
1. Отметка стимула (вспышка света); 2. Запись ЭЭГ затылочной области; 3. Сигнал ВП в процессе синхронного накопления (каждая последующая реакция складывается с предыдущими); 4. Маркер запуска, триггер; 5. Выделенный сигнал ВП с его параметрами.
 На рисунке 5 показано, как изменяется сигнал ВП и его соотношение с шумами ЭЭГ в процессе синхронного усреднения (рис.5.А). После первых 4 усреднений сигнал ВП еще не виден и скрыт в шумах спонтанной ритмики. После 9 и 16 усреднений мы его начинаем уже обнаруживать, но еще не очень четко. Только после 36 усреднений соотношение сигнала ВП к шуму спонтанной ритмики ЭЭГ достаточно, чтобы надежно отделить сигнал ВП от шума. Дальнейшее число усреднений улучшает соотношение сигнала к шуму, но уже в меньшей степени. На рис.5.Б показан пример спонтанной ритмики при синхронном усреднении без подачи стимула. После четырех усреднений уровень шума достигает 20 мкВ, при увеличении числа усреднений шум постепенно уменьшается («зануляется»). После 36 усреднений он составляет уже 8 мкВ, а после 100 равен 4 мкВ. При методике когерентного усреднения сигнал остается постоянным и, как бы постепенно, по мере увеличения числа усреднений, очищается от шума.
На рисунке 5 показано, как изменяется сигнал ВП и его соотношение с шумами ЭЭГ в процессе синхронного усреднения (рис.5.А). После первых 4 усреднений сигнал ВП еще не виден и скрыт в шумах спонтанной ритмики. После 9 и 16 усреднений мы его начинаем уже обнаруживать, но еще не очень четко. Только после 36 усреднений соотношение сигнала ВП к шуму спонтанной ритмики ЭЭГ достаточно, чтобы надежно отделить сигнал ВП от шума. Дальнейшее число усреднений улучшает соотношение сигнала к шуму, но уже в меньшей степени. На рис.5.Б показан пример спонтанной ритмики при синхронном усреднении без подачи стимула. После четырех усреднений уровень шума достигает 20 мкВ, при увеличении числа усреднений шум постепенно уменьшается («зануляется»). После 36 усреднений он составляет уже 8 мкВ, а после 100 равен 4 мкВ. При методике когерентного усреднения сигнал остается постоянным и, как бы постепенно, по мере увеличения числа усреднений, очищается от шума.
Рис.5. Выделение ВП методом пространственного синхронного усреднения на однократно предъявленную вспышку света.
A. ВП при последовательном их пространственном усреднении с 1-го по 16-ый электрод; Б. Схема расположения и обозначения электродов, с которых происходило пространственное усреднение; отведения монополярные с ипсилатеральным ушным индифферентным электродом; B. ВП, выделенный на одиночный стимул в результате пространственного усреднения реакций с 16 электродов. Эпоха анализа Т=500 мс; уровень сигнал/шум до выделения 0,84 (средний под разными электродами), после выделения 3,4 (по отношению к контрольной пространственной суммации без подачи стимула).
1. Инвазивные и неинвазивные методы исследования мозговых механизмов психических процессов.
2. Регистрации электрической активности мозга – электроэнцефалограмма (ЭЭГ).
3.Регистрация вызванных потенциалов (ВП) мозга и потенциалов, связанных с событиями.
4. Магнитоэнцефалография (МЭГ)
5.Томографические методы исследования мозга (компьютерная, позитронно-эмиссионная томография, метод ядерно-магнитного резонанса).
6. Электромиография.
7. Электроакулография.
8. Кожно-гальваническая реакция (КГР).
Темы рефератов:
1. История становления методов изучения мозга.
2. Полиграф.
3. История метода ВП.
4. Представления о механизмах генерации ВП мозга.
5. Основы принципа выделения ВП мозга.
6. Классификация ВП.
7. Принципы анализа ЭЭГ.
8. Основы метода ЭЭГ.
Литература:
Данилова Н.Н. Психофизиологическая диагностика функционального состояния. – М., 1992.
Данилова Н.Н. Психофизиология: Учебник для вузов. – М., 1998. – 378 с.
Гнездицкий В.В. Вызванные потенциалы мозга в клинической практике. – Таганрог., 1997. – 252 с.
Гречин В.б. Электроэнцефалография //Методы клинической нейрофизиологии. В серии: методы физиологических исследований. – Л., 1977. - Сс. 6-39.
Зенков Л.Р. Клиническая электроэнцефалография с элементами эпилептологии. _ Таганрог., 11996. – 358 с.
Лалянц И.Э., Милованова Л.С. томографы:психологические аспекты использования. – Вопросы психологии. – 1991. – Т.1. – СС. 143-148.
Магнитный резонанс в медицине /Под ред. П.А.Ринка. – М., 1995.
Наатанен Р. Внимание и функции мозга: учебное пособие/ Пер. с англ. Под ред. Е.Н.Соколова. – М.: Изд-во МГУ, 1998. – 560 с.
Новикова Л.А. Электроэнцефалография и ее использование для изучения функционального состояния мозга //Естественнонаучные основы психологии. – М., 1978.
Психофизиология. Учебник для вузов /Под ред. Ю.И.Александрова. – СПб., 2001. – 496 с.
Рутман Э.М. Вызванные потенциалы в психологии и психофизиологии. – М., 1975.
Русалова М.Н. Влияние информационного и эмоционального компонентов активации на ЭЭГ // Физиология человека. – 1980. – Т.6, № 3. – Сс. 410-419.
Тарханов И.Р. О гальванических явлениях в коже человека при раздражении органов чувств и различных формах психической деятельности //Вестник клинической и судебной психиатрии и невропатологии. – 1989. – Т. 7, № 1. – Сс. 73 – 81.
Трохачев А.И., Матвеев Ю.К. Регистрация активности нейронов головного мозга // Методы клинической нейрофизиологии / В серии: Методы физиологических исследований. – Л., 1977. – Сс.111-121.
Холодов Ю.А., Козлов А.Н., Горбач А.М. Магнитные поля биологических объектов. – М., 1987.