Механизмы питьевого поведения
Вода в живых организмах является основным растворителем, в котором происходит большинство химических реакций. За сутки взрослый человек с массой тела 70 кг теряет около 2,3 л воды: 1400 мл с мочой, 800 мл испаряется с поверхности кожи и уходит в виде водяных паров при дыхании, 100 мл – с калом. В то же время около 1 л воды поступает в организм с выпитыми жидкостями, примерно столько же содержится в съеденной пище и около 300 мл воды образуется при окислительных процессах в тканях – в сумме те же 2,3 л.
При уменьшении оптимального количества воды в организме возникает жажда. Она появляется, если потеря воды составит 0,5% от массы тела (при весе 70 кг ощущение жажды вызывает потеря всего лишь 350 мл жидкости). Утрата 10% от веса тела за счёт воды приводит к тяжёлой дегидратации (обезвоживанию), а потеря 20% является смертельной (10% = 7 л, которые теряются в норме за 2,5 суток).
Обмен воды неразрывно связан с обменом солей, а поэтому, говоря о воде, следует обсуждать проблему водно-солевого баланса. Растворённые в воде соли, в первую очередь натрия, создают осмотическое давление, которое заставляет воду перемещаться в область их большей концентрации. В организме вода распределена в трёх жидкостных пространствах: внутриклеточном, где содержится 2/3 всей воды, межклеточном или интерстициальном – приблизительно 1/4, а остальные 7-8% воды приходятся на плазму крови, лимфу и ликвор. Более высокая концентрация натрия во внеклеточной жидкости и крови, по сравнению с внутриклеточной, создаёт большее осмотическое давление. Оно уравновешивается онкотическим давлением в клетках, создаваемым растворёнными там белковыми молекулами. Таким образом, все жидкости оказываются изотоничными относительно друг друга.
Если концентрация натрия в крови и внеклеточной жидкости повысится, эти жидкости становятся гипертоничными (например, за счёт поступления большого количества соли с пищей или в связи с большой потерей воды при интенсивном потении). В таком случае вода начнёт перемещаться в область большей осмотической концентрации, т.е. из клеток во внеклеточное пространство. Потеря воды клетками (клеточная дегидратация), разумеется, крайне нежелательна, а устранить её можно, лишь восстановив прежнее, т.е. изотоничное состояние внеклеточной жидкости, – для этого надо просто разбавить её водой. Часть воды для этого можно сэкономить на диурезе, уменьшив на время объём выделяемой мочи, но одновременно необходимо увеличить количество выпиваемой жидкости.
Контроль величины осмотического давления осуществляют центральные и периферические осморецепторы. Функцию центральных рецепторов выполняют определённые нейроны супраоптических ядер гипоталамуса, периферические рецепторы находятся преимущественно в печени и воротной вене, несущей к печени кровь от пищеварительного тракта. Информация от периферических осморецепторов поступает к супраоптическим и паравентрикулярным ядрам гипоталамуса. При повышении осмотического давления нейроны гипоталамуса увеличивают секрецию антидиуретического гормона (АДГ, вазопрессин), который выделяется задней долей гипофиза.
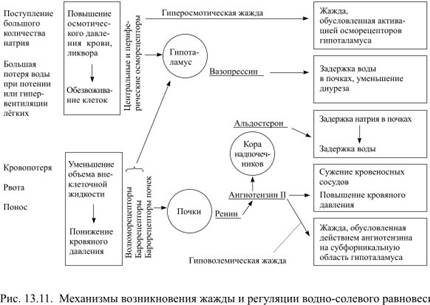 При уменьшении объёма внеклеточной жидкости и крови тоже возникает жажда, даже если осмотическое давление при этом не изменилось. Именно такая ситуация возникает после кровопотери, в том числе и у добровольных доноров. При уменьшении объёма циркулирующей крови меняется характер импульсации от волюморецепторов предсердий, а при падении кровяного давления ещё и от барорецепторов аорты и сонных артерий. Афферентная информация от этих рецепторов поступает в гипоталамус и стимулирует образование АДГ. Последующая задержка воды в почках способствует частичному восстановлению прежнего объёма внеклеточной жидкости.
При уменьшении объёма внеклеточной жидкости и крови тоже возникает жажда, даже если осмотическое давление при этом не изменилось. Именно такая ситуация возникает после кровопотери, в том числе и у добровольных доноров. При уменьшении объёма циркулирующей крови меняется характер импульсации от волюморецепторов предсердий, а при падении кровяного давления ещё и от барорецепторов аорты и сонных артерий. Афферентная информация от этих рецепторов поступает в гипоталамус и стимулирует образование АДГ. Последующая задержка воды в почках способствует частичному восстановлению прежнего объёма внеклеточной жидкости.
Уменьшение объёма крови приводит к резкому снижению кровоснабжения почек, которые обычно получают около 20% минутного объёма кровотока. В ответ на уменьшение кровотока почки выделяют в кровь ренин, а он, действуя как фермент на содержащийся в крови белок ангиотензиноген, способствует образованию из него ангиотензина. Появление ангиотензина сопровождается тремя важными последствиями: 1) он оказывает мощное сосудосуживающее действие, что исключительно важно при потере крови; 2) стимулирует образование АДГ в гипоталамусе и одновременно способствует появлению жажды; 3) повышает секрецию альдостерона корой надпочечников. Образующийся альдостерон увеличивает задержку ионов натрия в почках, а вслед за ионами натрия, подчиняясь механизму осмоса, задерживается вода, которая обычно следует за натрием, как нитка за иглой.
Конечно же, с помощью одних лишь механизмов гомеостатического регулирования восстановить потерянный объём жидкости, как и нарушенное осмотическое равновесие, невозможно без увеличения количества потребляемой жидкости. И нарушенное осмотическое равновесие, и уменьшение объёма внеклеточной воды вызывают жажду, которую в первом случае можно назвать гиперосмотической, а во втором – гиповолемической, но в обоих случаях питьевая мотивация возникает в связи с повышением активности нейронов гипоталамуса (рис. 13.11).
Главными из физиологических механизмов, способствующих окончанию питья, являются восстановление осмотического равновесия и/или потерянного объёма жидкости. Имеет значение и уменьшение сухости во рту, которую вызывало пониженное слюноотделение – неизбежное следствие экономии воды. Некоторое значение имеет само питьё, т.е. связанная с ним моторная деятельность и сенсорные ощущения. Определённую роль играет соответствующая сенсорная информация от рецепторов желудка и двенадцатиперстной кишки.
Многие люди, а может быть и большинство, склонны пить, ещё до того, как возникнет жажда: именно по этой причине питьё может просто сопровождать еду. Интересно, что в таком случае количество потребляемой жидкости зачастую пропорционально степени солёности пищи, т.е. на основе своего опыта человек может научиться предупреждать сдвиг водно-солевого равновесия. Количество выпиваемой жидкости нередко существенно зависит от её вкуса, от существующей традиции потребления определённых напитков (чай, квас, пиво и т.п.).