Мутации
1. Неопределенность (один и тот же внешний фактор может вызывать изменения разных признаков в разных направлениях, разные внешние факторы могут вызывать одинаковые изменения).
2. Степень изменения признака не зависит от силы или длительности действия внешнего фактора, вызвавшего изменение.
3. За редкими исключениями не имеют адаптивного значения.
4. Константны (не исчезают в течение жизни особи)
5. Наследуются
Наиболее важной областью практического приложения генетики является селекция — наука, разрабатывающая пути и методы создания новых и улучшения существующих форм домашних животных, возделываемых растений и полезных микроорганизмов. Генетика служит теоретической основой селекции: рационально планировать селекционные мероприятия можно лишь исходя из установленных генетикой общих законов наследственности и изменчивости и из более частных сведеннй о механизме наследования конкретных признаков, интересующих селекционера.
Количественные признаки. В предыдущих лекциях обсуждали наследование главным образом четко выражающихся в фенотипе признаков, относительно легко отличимых от альтернативных им, так что при расщеплении не возникает сомнения, к какому фенотипическому классу по данному признаку следует отнести ту или иную особь. Такие признаки (к ним относятся, например, особенности окраски или формы, наличие или отсутствие определенного фермента и т.п.) обычно называют качественными. Наряду с этим, многим наследственным признакам нельзя дать достаточно точного качественного описания, по ним между особями наблюдаются постепенные малозаметные переходы, а при расщеплении нет ясно разграниченных фенотипических классов. Эти признаки приходится изучать путем измерений или подсчетов, позволяющих дать признаку цифровую характеристику. К таким признакам принадлежат вес и размеры тела, плодовитость, урожайность, продуктивность, скороспелость, содержание белков и жиров, число одинаковых частей (например, зерен в колосе, лепестков цветка, кровяных клеток в определенном объеме) и т.п. Подобные признаки принято называть количественными.
Большинство хозяйственно-полезных признаков возделываемых растений и домашних животных относится к категории количественных, поэтому знание того, как наследуются количественные признаки, очень важно для селекционной работы. Для того, чтобы наследование количественных признаков стало понятным, нужно ознакомиться с некоторыми математически коконстантами, с помощью которых характеризуется значение таких признаков. Главные из этих констант — средняя арифметическая 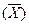 , среднее квадратическое отклонение (а), варианса (σ2),_ошибки средней арифметической (sх) и коэффициент изменчивости (CV). Вычисляются эти показатели следующим образом.
, среднее квадратическое отклонение (а), варианса (σ2),_ошибки средней арифметической (sх) и коэффициент изменчивости (CV). Вычисляются эти показатели следующим образом.
Прежде всего, изучаемый признак измеряют у всех особей исследуемой группы и полученные данные разбивают на произвольное, но относительно небольшое число классов (обычно на 10-15), в каждом из которых объединены особи, более или менее сходные по значению признака и которые составляют вариационный ряд.
Вариационный ряд удобно изображать графически в виде гистограммы- столбиков с основаниями, соответствующими классовому интервалу, и высотой, соответствующей числу вариант (т.е. измеренных особей) в классе, или в виде кривой распределения—из середины каждого класса восстанавливают перпендикуляр, высота которого соответствует числу вариант в классе, и вершины перпендикуляров соединяют прямыми линиями.
Количественные признаки, как правило, изменчивее качественных. Это зависит от двух причин. Во-первых, наследственные различия особей по тому или иному количественному признаку обусловлены обычно взаимодействием нескольких пар полимерных генов, причем каждый ген оказывает весьма существенное влияние на развитие данного признака; между тем, в случае качественных признаков различие между особями определяется, в основном, лишь одной, двумя или, изредка, тремя парами аллелей, а все прочие гены, влияющие на данный качественный признак, лишь в ограниченной степени могут модифицировать действие этих главных генов. Во-вторых, количественные признаки большей частью сильнее зависят от внешних факторов, чем качественные. Некоторые сочетания полимерных генов, определяющих количественный признак, сдвигают его значение в положительную сторону, другие — в отрицательную. Точно так же, разнонаправленно, действуют на фенотипическое выражение этих генов разные внешние влияния. В силу чисто статистических причин чаще всего встречаются особи, у которых значение количественного признака обусловлено приблизительно одинаковым суммарным действием благоприятных и неблагоприятных для развития этого признака независимых друг от друга наследственных факторов и факторов среды. Поэтому наиболее часты особи со средним значением количественного признака, а особи у которых это значение выше или ниже среднего, встречаются тем реже чем сильнее такое отклонение. Вследствие этой закономерности распределение частот особей с разным значением количественного признака обычно соответствует коэффициентам при отдельных членах разложения бинома Ньютона (а + b) при возведении его в разные степени:
(a+b)2 =a2+2ab+b2
(a+b)3=a3+3a2b+3ab2+b3
(a+b)4=a4+4a3b+6a2b2+4ab3+b4 и т.д.
откуда коэффициенты составляют такие ряды:
1+1
1+2+1
1+3+3+1
1+4 + 6+4+1 и т.д.
Теоретические кривые, построенные по коэффициентам разложения бинома при бесконечно большом числе вариант, называют биномиальными, а при бесконечно большом числе независимых факторов, положительно и отрицательно влияющих на значение признака, биномиальная кривая превращается в нормальную.
После того, как построен вариационный ряд, в первую очередь находят среднюю арифметическую , дающую представление о типичных размерах количественного признака в изучаемой группе особей.
При малом числе вариант (менее25) среднюю арифметическую вычисляют по формуле
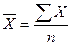
где — значение признака у отдельных особей, n - число особей, Σ —знак суммирования. При большом числе вариант этот способ слишком трудоемок и среднюю арифметическую вычисляют по формуле

где А — условная средняя; а — выраженное в числе классов отклонение каждого класса от класса, в котором находится условная средняя; f — число вариант в классе;  — выраженная в классах разность между условной средней и средней арифметической данного вариационного ряда; k - классовый интервал. В качестве условной средней обычно выбирают середину класса, содержащего наибольшее число вариант.
— выраженная в классах разность между условной средней и средней арифметической данного вариационного ряда; k - классовый интервал. В качестве условной средней обычно выбирают середину класса, содержащего наибольшее число вариант.
Между тем, вариационные ряды, имеющие одинаковую среднюю арифметическую, могут сильно различаться в отношении изменчивости признака. Наиболее удобным и употребительным мерилом изменчивости служит среднее квадратическое отклонение (σ). При малом числе вариант среднее квдратическое отклонение вычисляют по формуле
σ= 
если же в ряду более 25 вариант, то пользуются формулой
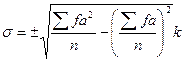
Квадратическое отклонение позволяет сравнивать изменчивость одного и того же признака в разных группах особей. Но, будучи числом именованным, этот показатель, очевидно не пригоден для сравнения изменчивости признаков, характеризуемых разными единицами измерения, например, изменчивости веса и роста и т. п. Для преодоления этой трудности обычно прибегают к вычислению коэффициента изменчивости (СV), представляющего собой выраженное в процентах частное от деления среднего квадратического отклонения на среднюю арифметическую: СV= 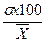 . Нужно, однако, подчеркнуть, что не следует пользоваться коэффициентом изменчивости там, где можно обойтись средним квадратическим отклонением — в математическом отношении последнее значительно точнее характеризует изменчивость признака.
. Нужно, однако, подчеркнуть, что не следует пользоваться коэффициентом изменчивости там, где можно обойтись средним квадратическим отклонением — в математическом отношении последнее значительно точнее характеризует изменчивость признака.
Любой эмпирический вариационный ряд представляет случайную выборку, содержащую ограниченное число вариант и поэтому в силу чисто статистических причин не вполне точно отражает характерную для данной популяции величину изучаемого признака и его изменчивость. Достоверность биометрических показателей, определяемых для эмпирической выборки, сильно зависит от числа составляющих ее особей — чем выборка многочисленнее, тем эти показатели ближе к своему истинному значению. При этом, биометрические показатели (средняя арифметическая, квадратическое отклонение, коэффициент изменчивости) отдельных эмпирических выборок распределяются вокруг истинных значений соответствующих показателей вполне определенным образом — в виде вариационнного ряда, близкого к нормальному; вариантами в этом ряду служат значения того или иного биометрического показателя, вычисленные для отдельных эмпирических выборок. Разброс выборочных показателей вокруг своих истинных средних можно измерить их средним квадратическим отклонением, т.е их σ. Среднее квадратическое отклонение биометрического показателя называется его средней квадратической ошибкой или, сокращенно, средней ошибкой(иногда говорят «стандартной ошибкой»). Средняя ошибка обозначается буквой S* со значком, указывающим к какому биометрическому показателю эта ошибка относится. Вычисление средних ошибок рассмотренных выше показателей ведется по следующим формулам:
средняя ошибка среднего квадратического
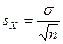 ;
;
средняя ошибка квадратического отклонения
 ;
; 
средняяошибка коэффициента изменчивости

Средние ошибки биометрических констант записывают со значком рядом со значением константы.
Наследование количественных признаков. При скрещивании особей, различающихся по количественному признаку, в F1, как правило, не наблюдается доминирования признака одного из родителей, а в F2 нет четкого расщепления на небольшое число фенотипически различных классов, находящихся между собою в определенных и характерных численных отношениях. Картина эта настолько отличается от типичной для наследования качественныхпризнаков, что в первые годы после вторичного открытия законов Менделя многим ученым, в том числе и таким крупным, как, например, Де Фризу, казалось невозможным распространять законы Менделя на наследование количественных признаков. Но довольно скоро выяснилось, что такая точка зрения ошибочна и что в основе наследственной передачи количественных признаков лежат те же явления расщепления и перекомбинации генов, что и при передаче качественных признаков.
В 1910 году шведский генетик Нильсон-Эле выдвинул предположение, согласно которому особенности наследования количественных признаков зависят от того, что признаки эти определяются полимерными генами. Правильность этого предположения в дальнейшем была подтверждена многими другими генетиками, изучавшими передачу разнообразных количественных признаков у разных растений и животных, и эта концепция, хотя и с некоторыми изменениями, лежит в основе современных представлений по данному вопросу.
Скрещивая краснозерную мягкую пшеницу с белозерной, Нильсон-Эле установил, что окраска зерен в F1, всегда промежуточна между родительскими и однородна, расщепление же в F2 различно в зависимости от того, какой краснозерный родитель участвовал в скрещивании. В некоторых случаях среди зерен F2 наблюдалось расщепление в отношениях 1очень светло-красное: 2светло-красным: 1 белому, в других — в отношениях темно-красное : 4 красным : 6 светло-красным : 4 очень светло-красным : 1 белому, в третьих же случаях расщепление было в отношении 63 окрашенных : 1 белому, причем цвет окрашенных зерен варьировал, так что среди них были все переходы от очень темно-красных до очень светло-красных. Эти результаты показали, что красная пигментация зерен развивается в присутствии любого из трех разных генов (обозначим их R1, R2 и R3), а белая окраска присуща зернам, гомозиготным по аллелям этих генов r1, r2 и r3 при этом все три гена красной окраски обладают примерно одинаковым по силе действием и действие этих генов может суммироваться. Таким образом, все белозерные растения имели генотип r1r1r2r2r3r3, а красные были гомозиготны либо по одному, либо по двум, либо по всем трем полимерным генам красной окраски. В первом случае скрещивание с белозерным растением дает в F2 моногибридное расщепление 1:2:1, во втором случае расщепление в F2 будет дигибридное. Отметим, что генотипически разные особи имеют здесь одинаковые фенотипы. В третьем случае расщепление будет тригибридным и, как нетрудно рассчитать, фенотипические классы F2 будут находиться в отношениях 1 :6: 15: 20 : 15 : 6: 1, т.е. зерна расположатся здесь по интенсивности окраски в биномиальный вариационный ряд, подобный наблюдаемым при наследовании других количественных признаков. Таким образом, особенности наследования количественных признаков вполне согласуются с законами Менделя, если исходить из представления, что такие признаки определяются полимерными генами.
Однако более или менее точное выяснение зависимости количественного признака от конкретных генов требует трудоемких исследований и к тому же возможно только у немногих генетически хорошо изученных организмов, у которых имеются линии с хромосомами, маркированными генами, определяющими удобные для наблюдения качественные признаки. Между тем, в практике растениеводства и животноводства постоянно возникает потребность решить, в какой мере изменчивость количественного признака в той или иной группе особей обусловлена генетическими причинами, а в какой— обязана модификациям, вызываемым окружающими условиями; от решения этого вопроса зависит прогноз эффективности отбора, проводимого по данному признаку. В таких случаях существенную пользу может принести вычисление коэффициента наследуемости изучаемого признака, что может быть сделано и без выяснения конкретных генов, определяющих этот признак.
Изменчивость всякого количественного признака в любой группе особей обусловлена совместным действием генетических факторов и разнообразных факторов окружающей среды (совокупность которых объединяют общим понятием паратипических). Коэффициент наследуемости представляет величину, показывающую, какова доля генетической компоненты в фенотипической изменчивости изучаемого количественного признака в рассматриваемой группе особей. Фенотипическая изменчивость признака характеризуется средним квадратическим отклонением или квадратом этого отклонения — вариансой (σ2). Общая фенотипическая варианса (σ2р) слагается из вариансы, зависящей от генетического разнообразия особей (σ2G), и вариансы, обусловленной паратипическими влияниями (σ2E). Отсюда следует, что доля генетической компоненты в общей изменчивости признака может быть выражена отношением  . Это и есть коэффициент наследуемости, обозначаемый h2. Ясно что разность между фенотипическими вариансами обеих групп будет приблизительно равна генотипической вариансе, т.е. σ2р2 - σ2р1 = σ2G, откуда легко вычислить коэффициент наследуемости изучаемого признака:
. Это и есть коэффициент наследуемости, обозначаемый h2. Ясно что разность между фенотипическими вариансами обеих групп будет приблизительно равна генотипической вариансе, т.е. σ2р2 - σ2р1 = σ2G, откуда легко вычислить коэффициент наследуемости изучаемого признака:

В селекционной практике вычисление коэффициента наследуемости имеет значение, главным образом, для прогнозирования эффективности отбора по тому или иному признаку: чем выше этот коэффициент в изучаемой группе : особей, тем больше шансов, что отборприведет у потомков к сдвигу признака в желаемую сторону. Например, у кур яйценоскость характеризуется низким, а вес яиц высоким коэффициентом наследуемости. Отбор был почти безрезультатным по яйценоскость и весьма успешным по весу яиц.
Системы скрещивания и их генетические следствия. Все системы скрещивания могут быть разделены по своим генетическим следствиям на два основных типа: скрещивания между родственными и между неродственными особями.
Инбридинг это скрещивание между родственными особями. В зависимости от степей родства скрещиваемых особей инбридинг может быть в разной мере тесным. Самый тесный из возможных видов инбридинга — самооплодотворение, широко распространенное среди растений, но встречаемое лишь у относительно немногих видов животных. У перекрестно оплодотворяющихся организмов наиболее тесный инбридинг — это скрещивания братьев с сестрами или родителей с детьми; несколько менее тесен инбридинг при скрещивании двоюродных братьев и сестер или племянников с дядьями и тетками; при более же отдаленном родстве скрещивающихся особей инбридинг становится соответственно еще менее тесным.
Первое важное генетическое следствие инбридинга состоит в повышении с каждым поколением гомозиготности потомков. Рассмотрим это на примере самого тесного инбридинга — самооплодотворения.
Самооплодотворение (в частности, самоопыление у растений) приводит к почти полной гомозиготизации уже в 10-м поколении.
Второе важное генетическое следстие инбридинга, тесно связанное с только что рассмотренным, заключается в разложении популяции на ряд генотипически различных линий. Такие линии тем многочисленнее, чем больше число генов, по которым гетерозиготна исходная популяция. В самом деле, при гетерозиготности ее по одному гену (Аа) возможно возникновение двух разных полностью гомозиготных линий (АА и аа), в случае гетерозиготпостн по двум генам (Аа Вb) их может быть четыре (ААВВ, ААаа, ааВВ и ааbb), когда же исходная популяция гетерозиготна по n-генам, то результатом инбридинга может быть выделение 2n различных гомозиготных линий. Вот почему инбридинг особей, взятых из панмиктической популяции, часто ведет к образованию большого числа генотипически различных. линий — ведь такие популяции обычно гетерозиготны по многим генам.
Инбридинг часто ведет к ослаблению и даже вырождению потомков.
Однако угнетающее действие инбридинга проявляется не всегда. Многие растения постоянно и без всяких признаков депрессии размножаются исключительно или преимущественно самоопылением, например, пшеница, овес, рис, горох, табак, томаты, ряд других возделываемых и множество диких видов растений. У лабораторных животных (мыши, крысы), у которых инбридинг большей частью приводит к вредным последствиям, в отдельных случаях даже очень длительный (50—70 поколений) тесный инбридинг не приводил к сколько-нибудь заметным отрицательным результатам. История создания некоторых высокопродуктивных пород домашних животных свидетельствует о том, что их родоначальниками были немногие особи, следовательно, эти в значительной степени инбридированы, что не отразилось на их достоинствах. У человека, у которого инбридинг, как правило, вреден, известны примеры длительного и тесного инбридинга, не сопровождавшегося вырождением. У перекрестнооплодотворяющихся организмов число предков всякой отдельной особи возрастает в каждом восходящем поколении в геометрической прогрессии. Так, в родительском поколении у нее 2 предка, в дедовском — 22 = 4 предка, в прадедовском 23 = 8 предков, а в n-м восходящем поколении число предков каждой особи равно 2n.Поскольку численность любого вида организмов в каждый данный момент представляет конечную величину, ясно, что число разныхпредков в каком-то более или менее далеком восходящем поколении должно было бы превысить численность вида, что невозможно. Отсюда вытекает, что среди предков происходили скрещивания между в той или иной степени родственными особями, вследствие чего фактическое число разных предков сокращалось.
Скрещивание между неродственными особями, или аутбридинг(outbreeding по-английски означает «неродственное разведение), по своим генетическим следствиям прямо противоположно инбридингу. Аутбридинг повышает гетерозиготность потомков, объединяет в гибридах аллели, существовавшие у родителей порознь. Вредные рецессивные гены, находившиеся у родителей в гомозиготном состоянии, подавляются у гетерозиготных по ним потомков, поэтому гибриды первого поколения часто оказываются более жизнеспособными, плодовитыми и продуктивными, чем родители, т.е. проявляют гетерозис(гибридную мощность).
Гетерозис.Явление гетерозиса (повышенной мощности гибридов первого поколения по сравнению с родительскими формами) известно давно. Впервые его описал еще в XVIII веке Кельрейтер на основании результатов своих опытов с растительными гибридами. Большое внимание уделял гетерозису Дарвин, собравший и подробно обсудивший множество примеров гетерозиса в своем труде о перекрестном опылении и самоопылении у растений. Первые важные генетические исследования гетерозиса были проведены в начале текущего столетия американскими генетиками Дж.Шеллом (ему принадлежит и сам термин гетерозис), Истом и Джонсом, а затем многие генетики разных стран вплоть до настоящего времени занимались изучением гетерозиса и разработкой способов его использования в растениеводстве и животноводстве. Выяснение природы гетерозиса пролило свет и на причины противоположного ему явления инбредной депрессии.
В итоге всех многочисленных исследовании в современной генетике сложилось представление, что гетерозис вызывается тремя основными причинами. Во-первых, это погашение у гетерозигот вредного действия рецессивных генов.
Во-вторых, в возникновении гетерозиса несомненно играет роль аддитивное действие благоприятных доминантных генов, присутствующих в разном наборе у родителей и соединяющихся в потомках, полученных скрещиванием этих родителей.
В третьих, гетерозис во многих случаях зависит частично, а иногда даже и целиком, от более полного или более благоприятного выражения некоторых генов в гетерозиготах, чем в гомозиготах. Если речь идет о доминантных генах с этим свойством, то говорят об их сверхдоминировании, однако лучше употреблять в применении к этому явлению термин моногибридный гетерозис,так как преимущество гетерозигот наблюдается не только в отношении некоторых доминантных, но и некоторых рецессивных генов.
Кроме этих трех основных причин гетерозиса (погашение у гибридов действия вредных рецессивов, суммация положительного влияния доминантных генов, полученных от обоих родителей и благоприятное проявление некоторыхгенов в гетерозиготном состоянии, известную роль в возникновении гетерозиса играют, по-видимому, и некоторые другие факторы, такие, как эпистатическое взаимодействие родительских генов в гибриде и взаимоотношения гибридного ядра с цитоплазмой происходящей только от матери (в частности, с ее митохондриями и пластидами), а у млекопитающих также иммунологические взаимоотношения матери и ее плода.
Получение гетерозисных гибридов прочно вошло в современную сельскохозяйственную практику. Особенно широко используется гетерозис при возделывании кукурузы, сорго, сахарной и кормовой свеклы, томатов и некоторых других овощей, а также при разведении свиней, кур и тутового шелкопряда. В ряде стран основные посевы кукурузы ведутся гетерозисными гибридными семенами и все промышленные выкормки тутового шелкопряда производятся из гетерозисной гибридной грены.
На примере кукурузы рассмотрим, как практически используется гетерозис. Сперва самоопылением создают много инбридированных (или инбредных,что то же самое) линий, добиваясь высокой степени их гомозиготности, о чем судят по их однородности в отношении морфологических и физиологических признаков; обычно это достигается после 4 — 6 поколений самоопыления. Затем эти линии скрещивают друг с другом во всевозможных сочетаниях, чтобы выявить их комбинационную способность,часто очень различную для разных таких сочетаний. После этого лучшие сочетания линий используют для получения гибридных семян. Однако такие семена образуются на относительно мало плодовитых инбредных родительских растениях и от каждого материнского растения удается собрать сравнительно немного семян. Поэтому семена для массовых посевов производятобычно с помощью двойной межлинейной гибридизации. Эти семена образуются на высокопродуктивных гетерозисных гибридных растениях, а развивающиеся из них растения сочетают гены четырех инбредных линий и по своим качествам не уступают гетерозисным растениям F1, получаемых двух инбредных линий.
Закрепить гетерозис удается там, где возможно вегетативное размножение гибридов — клубнями, луковицами, черенками. Так сохраняют положительный эффект гетерозиса, например, у картофеля, некоторых древесных и кустарниковых растений. В естественных условиях закрепление гетерозиса могло происходить при возникновении полиморфизма по инверсиям, например в природных популяциях дрозофил, или при установлении константной гетерозиготности по транслокациям, как это имело место у энотеры. Другой путь естественного закрепления гетерозиса — это переход к апогамии или другим формам бесполого размножения, что происходило в процессе эволюции многих видов растений. Гибридные растения часто превосходят родителей по активности ряда ферментов и богатству ростовых веществ.
Системы отбора.Отбор представляет собой важнейший элемент всякой селекционной работы.
Чтобы отбор был результативным, необходима информация о наследственном потенциале отбираемых особей. Такая информация с разной степенью полноты может быть получена тремя способами: изучением фенотипа особей, составляющих популяцию, в которой ведется отбор, анализом их родословных и исследованием оставляемого ими потомства.
Оценка фенотипа лежит в основе массового отбора.Массовый отбор стихийно применялся человечеством с незапамятных времен, затем сознательно использовался при выведении многих высокопродуктивных сортов культурных растений и пород домашних животных, не утратил своего значения и сейчас, так как он наиболее прост и доступен и в то же время нередко дает хорошие результаты. Но этот тип отбора имеет и существенные недостатки, обусловленные тем, что по фенотипу нельзя сколько-нибудь однозначно судить о генотипе отбираемых особей, а ведь только от их генотипа зависит эффективность отбора. Кроме того, в фенотипе не проявляются рецессивные гены, находящиеся в гетерозиготном состоянии, но могущие выявиться в потомстве. Поэтому массовый отбор, как правило, действует очень медленно, а иногда оказывается и вовсе безрезультатным.
Значительно совершеннее массового отбора индивидуальный отбор,использующий более полные сведения о генотипе отбираемых особей, чем могут дать наблюдения их фенотипа. При индивидуальном отборе популяцию делят на линии или семьи и проводят отбор отдельно в каждой из них, контролируя и учитывая всякое скрещивание. Критериями отбора, вдобавок к данным о фенотипе, служат сведения, доставляемые родословными, изучение которых позволяет судить о значении признаков у предков отбираемых особей, а также у их сибсов и полусибсов и других родичей. Кроме того, в некоторых случаях очень ценную информацию о генотипе этих особей удается получить, анализируя их потомство; этот способ оценки с большим успехом применяется в селекции животных.
При любых формах индивидуального отбора необходимо принимать во внимание, что успех зависит не только от правильного выбора скрещиваемых особей, но и от того, насколько благоприятно для потомства сочетание генов обоих родителей. Другими словами, в селекционной работе важен не только отбор, но и подборродительских пар по их генотипическим особенностям.
Закон гомологических рядов наследственнойизменчивости Н.И.Вавилова.Первым наиболее серьезным исследованием мутаций была работа Н.И.Вавилова по установлению параллелизма в наследственной изменчивости у видов растений, принадлежащих близким таксонам.
На базе обширных исследований морфологии различных рас растительного мира Вавилов в 1920 г. пришел к выводу, что, несмотря на резко выраженное разнообразие (полимофизм) многих видов, можно заметить ряд закономерностей в их изменчивости. Если взять для примера семейство злаков и рассмотреть варьирование некоторых признаков, то окажется, что одинаковые отклонения присущи всем видам.
Закон Вавилова гласит: «Виды и роды, генетически близкие, характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм у других видов и родов. Чем ближе генетически расположены в общей системе роды и линнеоны, т. е. виды, тем полнее сходство в рядах их изменчивости». Свой закон Н.И.Вавилов выразил формулой:
G1 (a + b + c …..)
G2 (a + b + c …..)
G3 (a + b + c …..)
где G1, G2, G3 — виды, а а, b, с - различные варьирующие признаки.
Для селекционной практики этот закон важен потому, что прогнозирует возможность найти неизвестные формы растений у данного вида, если они уже известны у других видов
Н.И.Вавилов положил закон гомологических рядов в наследственной изменчивости в основу поиска новых форм растений. Под его руководством были организованы многочисленные экспедиции по всему миру. Из разных стран были привезены сотни тысяч образцов семян культурных растений, составивших основу коллекций Всесоюзного института растениеводства (ВИР). Мутантные линии являются важнейшим исходным материалом при создании сортов культурных растений.