Гомеозисная мутация у дрозофил.
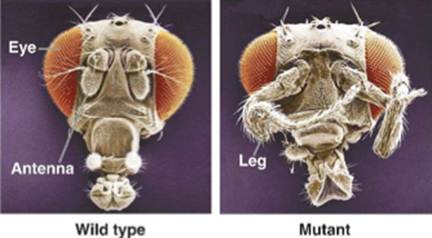
Трансформация передних разделов в задние из-за того, что у этого мутанта ген ANTP экспрессируется в голове, а не груди.
Ген Хокс с-6 контролирует образование грудного отдела позвоночника. У курицы ген Hoxc-6 отвечает за образование 7 грудных позвонков, образующих ребра. У змеи домен его экспрессии начинается от головы и простирается до клоаки → тело змеи = сплошная грудь.
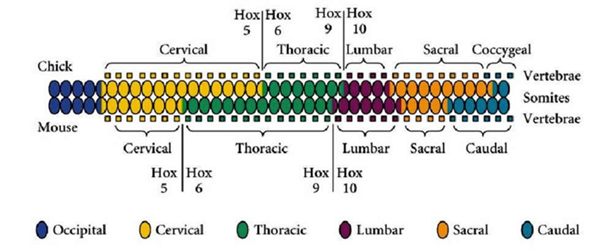
Сравнение экспрессии Hox-генов в курином и мышином эмбрионе выявляют различия в протяженности (но не порядке!) доменов экспрессии некоторых генов. Так, ген Hox 5 в длинной шейной области курицы экспрессировался в > чем 13 сомитах, а у мыши – в 6-7 сомитах. Т.е. НОХ-гены действительно детерминируют разные отделы тела эмбриона вдоль передне-задней оси!
Гомеобоксные хокс-гены считаются краеугольном камнем процессов развития у животных. Эдвард Льюис, Кристиа Нюсляйн-Фольхард и Эрик Вишаус за открытия, касающиеся генетического контроля на ранней стадии эмбрионального развития, получили Нобелевскую премию.
Хокс-гены включаются в онтогенез несколько раз. Они не только размечают участки тела вдоль передне-задней оси, но играют важную роль в развитии многих органов.Хокс-гены кластеров A и D размечают разные участки конечностей. О их роли в этом процессе свидетельствует фенотип мутантов (дефекты скелета конечностей).Мутации полидактилии выявлены у разных организмов. Фенотипическое сходство связано с мутациями в ортологичных НОХ-генах.
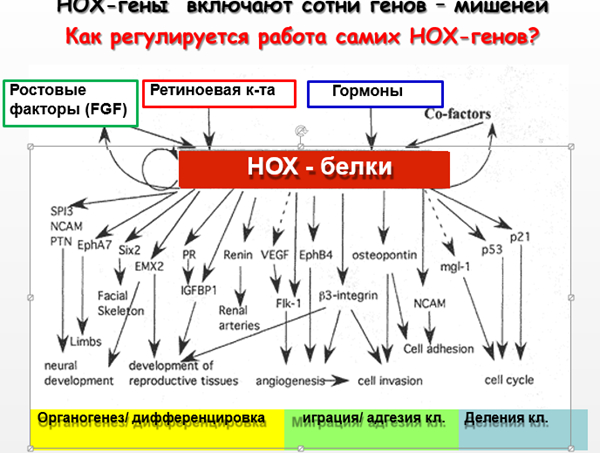
У хордовых и позвоночных в регуляции экспрессии генов важную роль играет ретиноевая кислота (производное от витамина А).
Обработка эмбрионов РК вызывает замещение средних отделов спинного мозга задними отделами.
При очень высоких концентрациях наблюдалось недоразвитие задних отделов эмбриона. Это связано с регуляцией РК-экспрессии Хокс-генов. Алкоголь через РК влияет на экспрессию многих НОХ-генов, которые важны не только на самых ранних этапах эмбриогенеза, но и на стадии гаструляции и органогенеза. Изменения экспрессии регуляторных генов, контролирующих развитие, приводят к нарушению развития (уродства). Однако эти нарушения, как правило, не наследуются, т.к. не связаны с мутациями (модификации). Тем не менее есть и особый тип модификаций, которые могут передаваться через деления клеток и даже сохраняться потомками. Это эпигенетические изменения (эпи – над), т.е. длительно сохраняющиеся изменения экспрессии генов, не связанные с изменениями последовательностей ДНК и РНК. Именно эти механизмы используются для того, чтобы вызвать молчание 90% генов в каждом типе специализированных клеток.
Эпигенетические изменения связаны главным образом с процессами метилирования ДНК и гистоновых белков, которое осуществляется специальными ферментами. Это взаимосвязанные процессы. Ферменты работают в составе сложных белковых комплексов. Главное отличие эпигенетических механизмов регуляции работы генов от других механизмов, которые используются в регуляции развития, заключается в долговременности их действия, что создает впечатление закрепления изменений на уровне генетического кода. Эпигенетические механизмы объясняют многие удивительные феномены. Кастовая структура общественных насекомых также связана с эпигенетическими механизмами. У пчел матка (царица), нянька и сборщица меда имеют одинаковый генотип, но разный эпигенотип, что связано с характером кормления.
Внешняя среда вступает в диалог с геном уже во время внутриутробного развития. Монозиготные близнецы имеют одинаковый генотип, но у каждого есть своя пуповина и плацента. Микроусловия разные → различие эпигенетических меток (это механизм адаптации к условиям среды). Именно эпигенетическими изменениями объясняют те различия между монозиготными близнецами, которые изредка все же встречаются, несмотря на то, что они имеют одинаковый генотип (их эпигенотип может отличаться!).
Наш образ жизни может помешать реализации генетических программ развития у детей и внуков.