Протеазы (пептидгидролазы)
Белок и пептиды расщепляют ферменты, объединяемые в подкласс пептидгидролаз (К.Ф.3.4.). Их называют также протеазами, протеолитическими ферментами.
Основной реакцией, катализируемой протеолитическими ферментами, является гидролиз пептидной связи в молекулах белков и пептидов:
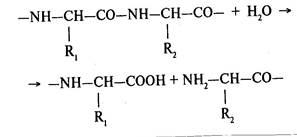
Расщепление пептидной связи в точках, удаленных от конца молекулы, катализируют эндопептидазы. Их делят на четыре группы: сериновые (К.Ф.3.4.21), тиоловые (К.Ф.3.4.22), карбоксильные (К.Ф.3.4.23) и металлосодержащие (К.Ф.3.4.24).
Отщепление концевых аминокислот и дипептидов, а также гидролиз дипептидов катализируют экзопептидазы, которые разделяют на пять групп. Аминопептидазы (К.Ф.3.4.11) катализируют отщепление единичных аминокислот от N-конца полипептидной цепи, карбоксипептидазы (К.Ф.3.4.16, 3.4.17) – от С-конца. Дипептидазы (К.Ф.3.4.13) гидролизуют дипептиды. Дипептидилпептидазы (К.Ф.3.4.14) и пептидилдипептидазы (К.Ф.3.4.15) катализируют отщепление дипептидов соответственно от N-конца и от С-конца полипептидной цепи. Схема классификации дана на рис 7.1
 |
Существующая классификация пептидгидролаз несовершенна, поскольку в ней используются в качестве разграничительных различные признаки: деление пептидаз на группы проведено по характеру действия на субстрат, а протеиназ – по структуре каталитического центра. В настоящее время описано несколько сот пептидгидролаз различного происхождения. На основании этого обширного материала предлагаются новые варианты классификации, в частности, только по структуре каталитического центра.
Скорость ферментативного гидролиза белковых соединений определяется наличием в них пептидных связей, специфичных для действия фермента, а также пространственной структурой субстрата.
На доступность пептидных связей гидролизу влияют вторичная, третичная и четвертичная структура белков. Белки могут иметь один или два типа упорядоченных вторичных структур (α-спиральной и β-складчатой), представленных в различных сочетаниях и охватывающих более или менее значительную часть полипептидной цепи.
В упорядоченных структурах определенные участки полипептидной цепи экранированы и недоступны действию ферментов. Чем выше степень упорядоченности структуры, тем менее белок подвержен протеолизу.
Третичная структура белка, его геометрическая форма определяет соотношение экспонированной и экранированной частей молекулы (то есть доступной и недоступной протеолизу),
Наименее доступны протеолизу молекулы с наименьшей удельной поверхностью, то есть приближающиеся по форме к шару.
Четвертичную структуру имеют белковые молекулы, состоящие из субъединиц. Последние могут быть ассоциированы за счет ковалентных, ионных и водородных связей. Ассоциация субъединиц снижает относительную величину экспонированной части молекулы, увеличивает ее конформационную стабильность за счет внутримолекулярных взаимодействий. Ассоциированные молекулы менее доступны действию ферментов, чем диссоциированные.
Денатурация белков сопровождается развертыванием полипептидной цепи, демаскированием прежде экранированных групп. Снимаются ограничения доступности субстрата, обусловленные вторичной, третичной четвертичной структурой. Денатурированные белки гидролизуются, целом быстрее и полнее нативных.
Сериновые протеиназы объединяют группу эндопептидаз, имеющих в каталитическом центре остаток серина. У большинства сериновых протеиназ в каталитическом центре находят аминокислотные остатки Асп 102, Гис 57 и Сер 195. Сериновыё протеазы ингибируют агентами, взаимодействующими с остатком серина, такими как диизопропилфторфосфат (DFP), фенилметилсульфонилфторид (PMSF), сульфогалогениды, а также природными протеолитическими ингибитора животного, растительного и микробного происхождения. Все сериновые протеиназы проявляют максимальную активность в нейтральной или щелочной среде. Молекулярная масса составляет от 12-15 кДа у ферментов с единственной полипептидной цепью, до 200-300 кДа – у ферментов с субъединичной структурой.
К группе сериновых протеиназ относятся ферменты животного, растительного и микробного происхождения, многие из которых хорошо изучены и выпускаются в виде промышленных препаратов.
В поджелудочной железе человека и млекопитающих животных синтезируются сериновыё протеиназы – трипсин (К.Ф.3.4.21.4), химотрипсины А и В (К.Ф.3.4.21.1), химотрипсин С (К.Ф.3.4.21.2). Эти ферменты образуются первоначально в виде предшественников (трипсиногена и химотрипсиногенов), которые превращаются в активные формы под действием трипсина. При протеолитическом процессинге трипсиногена от его молекулы отщепляется N-концевой гексапептид, реакция активируется кальцием.
Активная форма трипсина – однокомпонентный белок, состоящий из единственной полипептидной цепи. Фермент, выделенный из поджелудочной железы человека и различных млекопитающих, имеет молекулярную массу 22,9-25,5 кДа, оптимальный рН 7-8. Стабилен при рН не выше 6. В щелочной среде происходит самопереваривание. В организме трипсин стабилизируется с помощью ингибитора, с которым образует неактивный комплекс, устойчивый к протеолизу. Ингибитор синтезируется в поджелудочной железе. Трипсин гидролизует пептидные связи, образованные карбоксилами аргинина или лизина. Существенно наличие свободной аминогруппы диаминокислот по соседству с расщепляемой связью. Фермент гидролизует не только амидные, но также сложноэфирные связи. Он расщепляет различные животные и растительные белки, проявляет высокую активность в отношении белковых компонентов клеточных стенок микроорганизмов. Комплекс трипсина и литической мурамидазы – лизоцима, секретируемого слизистой кишечника, является важнейшим фактором регуляции состава микрофлоры пищеварительного тракта животных и человека.
Химотрипсины образуются из химотрипсиногенов после отщепления пептида, состоящего из 15 аминокислотных остатков. Молекулярная масса химотрипсинов составляет 24-25,8 кДа, оптимальный рН 7-9. Химотрипсины стабильны в кислой среде. Ферменты предпочтительно катализируют расщепление в белках пептидных связей, в образовании которых участвуют карбоксилы ароматических аминокислот. Гидролизуют также связи, образованные карбоксилами лейцина, метионина, триптофана. Химотрипсины, как и трипсин, расщепляют сложные эфиры. Разнообразие типов гидролизуемых связей определяет широкую специфичность действия химотрипсинов.
В группу сериновых входят многие внеклеточные и внутриклеточные протеиназы микроорганизмов. Классическими представителями секретируемых сериновых протеиназ бактерий являются субтилизины А, В и BPN' из культуры В. subtilis – белки молекулярной массы 26,3-27,5 кДа с оптимумом действия при рН 8-11. Многие бациллы продуцируют субтилизиноподобные сериновыё протеиназы.
Для внеклеточных сериновых протеиназ бактерий характерен оптимум действия в щелочной зоне (рН 8-12,5), при температуре 50-70° С. Ферменты стабильны в зоне рН от 6 до 13, при температуре не выше 50° С. В качестве стабилизатора могут использоваться соли кальция.
Сериновыё протеиназы входят в состав комплексных ферментных препаратов, выделяемых из грибов аспергиллов. У Asp. oryzae найдены разновидности сериновых протеиназ, которые при действии на казеин проявляют максимальную активность при рН 7-8,5 и 9-11,5.
Многие виды микроскопических грибов (p. p. Acremonium, Alternaria, Aspergillus, Cephalosporium, Fusarium, Penicillium, Trichoderma, Verticillium и др.) синтезируют внеклеточные сериновыё протеиназы. Большинство исследованных ферментов этой группы имеют молекулярную массу 18-35 кДа, оптимум действия – при рН от 5 до 12 и температуре 55-60° С. Многие сериновыё протеиназы грибов стабилизируются ионами кальция.
В соке растений присутствуют растворимые формы субтилизиноподобных сериновых протеиназ. Их выделили из плодов дыни, различных видов тыквы, из томатов, плодов маклюры (сем. тутовых), из листьев подсолнечника, корней одуванчика, клубеньков ольхи. Все исследованные растительные субтилизины – гликопротеины, фермент из маклюры содержит 25,6% углеводов. Они синтезируются в виде препроферментов, которые после протеолитического процессинга превращаются в активные формы. Молекулярная масса активных форм - 50-70 кДа, у подсолнечника – 25 кДа. Оптимальный рН действия – в интервале 7,3-11, при температуре 55-70° С. Стабильны при температуре не выше 50-60° С. Растительные субтилизины имеют широкую зону рН-стабильности: фермент из тыквы – 5-10, дыни – 4-12, подсолнечника – 4-10. Гидролизуют казеин, азоказеин, азоколлаген, гемоглобин и другие белки, преимущественно расщепляя связи, образованные гидрофобными аминокислотами (лейцином, фенилаланином, тирозином), с меньшей скоростью – образованные цистеином, дикарбоновыми аминокислотами и их амидами.
Растения синтезируют различные ингибиторы ферментов, среди которых наиболее широко представлены ингибиторы сериновых протеиназ, в частности, ингибиторы трипсина. Они найдены в семенах бобовых (сои, фасоли, гороха), злаков (пшеницы, ржи, кукурузы), гречихи, в клубнях картофеля, корнеплодах моркови, яблоках, листьях капусты и других источниках. Ингибиторы трипсина предотвращают инфицирование растений микроорганизмами и повреждение насекомыми, поскольку большая часть патогенных микроорганизмов и насекомых-вредителей синтезирует трипсиноподобные ферменты. В семенах гречихи найден медленно взаимодействующий и прочно связывающийся ингибитор трипсина, подавляющий прорастание спор и рост мицелия патогенного гриба Alternaria alternata. В клубнях картофеля присутствуют две разновидности ингибиторов сериновых протеиназ с молекулярной массой 21 и 22 кДа. Первый подавляет активность трипсина, химотрипсина и эластазы, второй – только двух первых ферментов. Среди ингибиторов химотрипсина, синтезирующихся в тканях картофеля, нашли высокоэффективный пептид массы всего 5 кДа. Расшифровка структуры низкомолекулярного ингибитора перспективна в целях создания синтетических аналогов для применения в качестве средств защиты растений и медицинских препаратов.
В семенах сои – две разновидности ингибиторов трипсина: ингибитор Кунитца и ингибитор Баумана-Бирк (ББИ). Второй способен одновременно и независимо связывать на одном центре – трипсин, а на другом – эластазу лейкоцитов человека и химотрипсиноподобные ферменты, за что получил название «двухголового». Показано, что ББИ обладает антиканцерогенным действием, препятствуя метастазированию и росту злокачественных опухолей, происходящему при участии лейкоцитарных ферментов химотрипсинового типа. Содержание ББИ в разных сортах сои составляет от 0,5 до 1,5 мг/г семян, в некоторых видах коммерческой соевой муки – до 1,9 мг/г. В семенах сои ББИ представлен несколькими изоформами, число которых, в зависимости от сорта сои, составляет от 2 до 7. Высокая биологическая активность ББИ побудила пересмотреть выводы об антипитательных свойствах соевых ингибиторов. Считают, что антипитательные свойства связаны с присутствием в сое большого количества таннинов – неспецифических ферментных ингибиторов широкого спектра, а также гемаглютининов.
Помимо ингибиторов собственно сериновых протеиназ, растения синтезируют бифункциональные ингибиторы, подавляющие активность α-амилазы и сериновых протеиназ. Такие ингибиторы характерны для семян злаков. В процессах гидролиза растительных белков необходимо принимать во внимание присутствие ингибиторов в растительном сырье.
Сериновыё протеиназы животного и микробного происхождения прочно вошли в номенклатуру промышленных феррментных препаратов. Из животного сырья производят трипсин, химотрипсин и комплексный препарат панкреатин с активностью обоих этих ферментов. Из микробного сырья получают Проназу, Римопротелин, Протосубтилин щелочной, Амилопроторизин, Лизосубтилин и другие препараты.
Тиоловые (цистеиновые) протеиназы. В эту группу входят эндопептидазы, для проявления каталитической активности которых существенна сульфгидрильная группа цистеина. Тиоловые протеиназы ингибируются парахлормеркурибензоатом (PCMB), монойодуксусной кислотой (MIA) и другими окислителями, а также ионами тяжелых металлов. Активируются восстановителями (2-меркаптоэтанолом, дитиотреитолом, цистеином, бисульфитом натрия и пр.).
Большинство тиоловых протеиназ имеют рН-оптимум в слабокислой или нейтральной зоне. К тиоловым принадлежат широко применяемые в практике протеиназы растительного происхождения – папаин, химопапаины А и В, бромелаин, фицин.
Папаин (К.Ф.3.4.22.2) и химопапаины А и В (К.Ф.3.4.22.6) выделяют из млечного сока дынного дерева. Это ферменты молекулярной массы 23,4-28 кДа с оптимумом рН 6-7. Папаин стабилен в зоне рН 3-11. Наибольшая стабильность наблюдается при рН 5. В присутствии окислителей папаин быстро теряет активность, поэтому при работе с ним необходимо следить за окислительным потенциалом среды.
Бромелаин (К.Ф.3.4.22.4) выделяют из сока зрелых стеблей ананаса, фермент имеет массу 35 кДа, оптимальный рН 5-6.
Фицин (К.Ф.3.4.22.3) получаемый из латекса тропического инжира, имеет молекулярную массу 23,8кДа и оптимальный рН 6,5-9,5.
Папаин, бромелаин, фицин - протеиназы широкой субстратной специфичности. Они гидролизуют в белках пептидные связи, образованные лейцином или глицином. Другие типы связей, в том числе характерные для специфичности пепсина, трипсина, химотрипсина, карбоксипептидазы, также могут расщепляться, хотя и с меньшей скоростью. Благодаря этому растительные тиоловые протеиназы более глубоко гидролизуют белки, чем сериновыё и карбоксильные протеиназы животного происхождения.
В растениях найдены ингибиторы цистеиновых протеиназ. Из клубней картофеля изолирован белок молекулярной массы 23 кДа, подавляющий активность папаина, бромелаина и фицина. В семенах тыквы – ингибитор молекулярной массы 7,5 кДа, эффективно подавляющий фицин и папаин, менее – химопапаин, на бромелаин не действует. В семенах сои содержатся две формы ингибитора цистеиновых протеиназ основная, с молекулярной массой 14 кДа, и минорная – 8,2 кДа. Основной компонент ингибирует папаин, бромелаин, фицин, химопапаин. В его молекуле три дисульфидных моста, разрыв которых под действием восстановителей (цистеина, цистамина) приводит к обратимой инактивации. Окислители реактивируют ингибитор.
Тиоловые протеиназы продуцируют эубактерии (клостридии, стрептококки, стафилококки) и архебактерии. У грибов их находят редко. Микробные тиоловые протеиназы наиболее активны при рН 7-8,8.
Карбоксильные протеиназы распространены у животных и эукариотических микроорганизмов. В каталитическом центре карбоксильных протеиназ – две карбоксильные группы. Активность подавляют эпоксисоединения, диазокетоны, природные ингибиторы типа пепстатина.
Карбоксильные, или кислые, протеиназы, за малыми исключениями, проявляют активность в кислой среде.
К числу кислых протеиназ относятся пепсины млекопитающих пепсин А (К.Ф.3.4.23.1), пепсин В (К.Ф.3.4.23.2) и пепсин С (К.Ф.3.4.23.3). Это белки молекулярной массы 31-40 кДа с оптимумом действия при рН2-4.
В слизистой желудка человека синтезируется предшественник пепсина – пепсиноген (42 кДа), который превращается в пепсин (34-35 кДа)| в присутствии кислоты или автокаталитически, в результате гидролиза активным пепсином.
Пепсин проявляет специфичность в отношении связей, образованных аминофуппами тирозина или фенилаланина. Наличие свободной аминофупиы вблизи пептидной связи препятствует гидролизу белка, а карбоксильной – увеличивает скорость гидролиза. Пепсин расщепляет различные животные и растительные белки, в том числе казеин, коллаген, глютин, эластин, кератин, гистон.
Способность синтезировать кислые протеиназы широко распространена у микроскопических грибов – аспергиллов, пенициллов, мукоров, Rhizopus, Trichoderma и др. Внеклеточные кислые протеиназы грибов – ферменты молекулярной массы от 30 до 100 кДа (чаще 32-39 кДа), с оптимумом действия при рН 2-4,6. Стабильны в кислой среде (рН 2,5-6), в нейтральной среде быстро и необратимо инактивируются. Оптимальная температура действия 55-75°С, большинство исследованных кислых протеиназ устойчивы при температуре до 55° С, некоторые при 60-65° С. По специфичности действия кислые протеиназы грибов сходны с пепсином.
Внеклеточные кислые протеиназы грибов, наряду с другими видами протеолитических ферментов, входят в состав комплексных ферментных препаратов, таких как Амилоризин, Амилопроторизин, Пектаваморин, Пектофоетидин, Целловиридин, Целлюлаза-100 и др. Из аспергиллов выделяют очищенные препараты кислойпротеиназы, предназначенные для пищевой промышленности и медицины.
Металлосодержащие протеиназы – это ферменты, в каталитический центр которых входит ион или ионы металлов, существенные для проявления каталитической активности. Такими металлами чаще всего являются цинк, кобальт или марганец. Некоторые металлопротеиназы содержат как единственный металл кальций или магний. Кальций может входить в молекулу фермента также как ион-стабилизатор. Так в металлопротеиназе В. subtilis цинк является каталитически активным металлом, а кальций – стабилизатором.
Металлы играют важную роль в каталитическом акте. С их помощью образуется тройной координационный комплекс: фермент-металл-субстрат. При этом стабилизируется высшее валентное состояние металла. Ионы металлов могут образовывать координационные связи с атомами азота, серы, кислорода, имеющими неподеленные пары электронов. Отток электронов от групп >С=0 и -NH- к иону металла облегчает разрыв пептидной связи.
Все металлопротеиназы ингибируются хелатными агентами (EDTA, EGTA, о-фенантролином и др.). Инактивация чаще всего носит обратимый характер.
Металлопротеиназы широко распространены у бактерий встречаются и у микроскопических грибов. Бактериальные металлопротеиназы – белки с молекулярной массой 28-48 кДа, проявляющие максимальную активность при рН 6,5-9 и температуре 50-75° С.
В. subtilis – продуцент протеолитического ферментного препарата Протосубтилина выделяет в среду две металлопротеиназы – А и Б, с молекулярной массой 44 и 40 кДа. Первая имеет оптимум при рН 7, вторая – при рН 7,5 и 11.
Микроскопические грибы синтезируют металлопротеиназы молекулярной массы 20–45 кДа. Среди них выделяют группу кислых ферментов, с оптимальным рН 5-6, и нейтральных, с оптимумом рН около 7 Температурный оптимум лежит обычно в пределах 45-55 С, у металлопротеиназы Asp. sojae – при 650С.
Среди металлопротеиназ аспергиллов и пенициллов находят цинксодержащие ферменты (1 ион цинка на молекулу).
Карбоксипептидазы широко представлены у животных, растений и микроорганизмов, они играют важную роль в белковом обмене. Среди карбоксипептидаз встречаются сериновыё и металлоферменты, редко – тиоловые.
В поджелудочной железе синтезируются карбоксипептидазы А и В. Предшественники этих ферментов активируются под действием трипсина. Карбоксипептидаза А – металлофермент, содержит один атом цинка на молекулу
Молекулярная масса фермента 34,3 кДа, оптимальный рН 7,5, зона стабильности – рН 6-10,2. Карбоксипептидаза В идентична по массе и содержанию цинка, стабильна при рН 7-9. Оба фермента имеют как пептидазную, так и эстеразную активность.
Карбоксипептидазы есть у многих покрытосеменных (томата, арбуза, фасоли, шпината и др.). В клетках растений присутствуют, как связанные, так и растворимые формы карбоксипептидаз. Максимальную активность находят во фракции митохондрий.
Все растительные карбоксипептидазы – кислые белки с изоэлектрической точкой при рН 4,3-5,4. Молекулярная масса 90-175 кДа. Оптимальный рН 5-5,6, стабильны в зоне рН 4-6. Оптимальная температура действия 40-50° С.
Большинство растительных карбоксипептидаз относится к сериновым ферментам. Из их числа – уникальная карбоксипептидаза фасоли, способная отщеплять с С-концов белков все аминокислоты, за исключением аспарагиновой. Предпочтительно наличие на С-конце аминокислоты с длинной алифатической цепью, а по соседству с ней – ароматаческой аминокислоты.
Карбоксипептидаза из кожуры апельсина является металлоферментом, содержащим 3,6 атома цинка на молекулу. Фермент отщепляет с С-конца белков ароматические, кислые аминокислоты и пролин. Приведенные примеры свидетельствуют о широкой субстратной специфичности растительных карбоксипептидаз.
Микробные карбоксипептидазы – белки молекулярной массы от 30 до 160 кДа. Оптимальный рН действия для грибных карбоксипептидаз – 3,0-6,8, для бактериальных - 8,0-9,8. Оптимальная температура действия карбоксипептидаз Asp. oryzae - 40-50° С.
Карбоксипептидазы микроорганизмов могут иметь различную структуру активного центра. У некоторых грибов и актиномицетов они являются металлоферментами, для проявления активности которых необходимы ионы цинка, марганца, магния, кальция.
Карбоксипептидазы микроскопических грибов характеризуются широкой субстратной специфичностью, что в значительной мере определяет способность грибных ферментных препаратов катализировать глубокое расщепление растительных и животных белков. Карбоксипептидазы пенициллов отщепляют от пептидов остатки пролина, лизина, аргинина, глицина, дикарбоновых аминокислот. Кислая карбоксипептидаза Asp. saitoi последовательно отщепляет все аминокислоты от нативного инсулина и некоторых других пептидов. Аналогичный фермент Asp. sojae глубоко расщепляет соевый белок.
Протеолитические ферменты семян растений. В семенах злаковых и бобовых культур содержится целый комплекс протеолитических ферментов, участвующих в расщеплении запасных белков до аминокислот в процессе прорастания семян.
В настоящее время известно, что протеолиз белков в семенах растений осуществляется комплексом ферментов, различающихся по своим функциям, механизму действия и другим показателям. Некоторые из этих ферментов были выделены в виде высокоочищенных препаратов и подробно охарактеризованы. Так, например, из семян пшеницы были выделены несколько типов протеолитических ферментов, различающихся по оптимуму рН: кислые протеиназы с оптимумом рН 3,7-4,0; нейтральные протеиназы с оптимумом рН 6,5-7,0; щелочные протеиназы с оптимумом рН > 8,0.
Из трех групп протеиназ наибольшего внимания технологов заслуживают нейтральные протеиназы. По активности они в несколько раз превосходят кислые и в условиях теста способны эффективно расщеплять белки клейковины. Одна из особенностей нейтральных протеиназ состоит в том, что они не растворяются в водных, солевых и буферных растворах. Они прочно связаны с белками клейковинного комплекса и извлекаются при частичном растворении клейковины в щелочном растворе. Максимальное извлечение нейтральных протеиназ происходит при обработке измельченного зерна, муки или лиофилизированной клейковины 0,35%-м раствором карбоната натрия созревших семян пшеницы нейтральные протеиназы и их белковые ингибиторы образуют единый неактивный комплекс, связанный с клейковиной. Соотношение активности протеиназ и их ингибиторов в созревшем зерне определяет стабильность белкового комплекса, его устойчивости в процессе тестирования.
Нейтральные протеиназы не активируются восстановленным глута-тионом или цистеином и поэтому не могут быть отнесены к тиоловым ферментам, в отличие от кислых протеиназ. Нейтральные протеиназы ингибируются хлоридом натрия, фенольными соединениями, фенольными соединениями, ароматическими аминокислотами, продуктами сахаро-аминной реакции (меланоидинами).
Хлорид натрия является обязательным компонентов рецептуры и, внесенный в таком количестве, снижает активность нейтральных протеиназ и соответственно интенсивность автолиза на 60-70%.
При переработке слабой муки необходимо как можно раньше вводить соль, тогда как для муки с чрезмерно крепкой клейковиной желательно активизировать протеолиз, и соль следует вносить на более поздних стадиях.
В связи с этим необходимо еще раз подчеркнуть важность изучения собственных эндогенных ферментных систем биологического сырья, факторов, влияющих на их активность с точки зрения их огромной роли в процессах, происходящих при созревании, хранении и переработке пищевого сырья.