Популяции в экосистеме. Основы демографии
Под популяцией можно понимать любую достаточно обособленную группу организмов одного вида, занимающую определенное пространство и функционирующую как часть экосистемы.
Различные виды организмов представлены в экосистеме именно популяциями. Поэтому популяцию можно рассматривать как систему, стоящую на иерархической лестнице природных систем между уровнем отдельных организмов и уровнем экосистем. Экосистемы могут существовать относительно обособленно, как отдельные организмы, например экосистемы островов, а популяции в изолированном виде нигде не встречаются.
Для человека знание законов популяционной динамики имеет одно из первостепенных значений, так как эти законы применимы и к динамике человеческих популяций. Особую важность эти знания приобретают в связи с демографической проблемой, являющейся одним из основных компонентов надвигающейся глобальной экологической катастрофы. Однако человек в некоторой степени обособился от природы и обладает мощными средствами воздействия на среду. Поэтому, например, неправильными оказались прогнозы демографов прошлого (Томас Мальтус), регулярно предсказывавших нам скорую стабилизацию численности населения планеты, чего до сих пор не произошло.
1.5.1. Параметры и закономерности динамики популяций
Рассмотрим ряд свойств, которыми характеризуется популяция. Эти свойства можно разделить на биологические и групповые. Биологические свойства присущи как популяции в целом, так и составляющим ее организмам. Они характеризуют жизненный цикл популяции: популяция, так же как и отдельный организм, растет и поддерживает сама себя, имеет определенную организацию и структуру. Групповые свойства присущи только группе в целом. Как правило, это статистические параметры, такие как плотность, рождаемость, смертность, возрастная структура. В числе прочего эти параметры характеризуют динамику популяции, то есть изменение во времени ее численности, структуры и т.п.
1.5.1.1. Статистические параметры
Плотность популяции выражается количеством особей или биомассой на единице пространства, например число деревьев на 1 га, или масса планктона в 1 м3. На разных стадиях жизненного цикла плотность может существенно колебаться. Это напрямую связано с двумя другими показателями популяции: рождаемость и смертность.
Рождаемость DNn - это способность популяции к увеличению численности, независимо от того, происходит ли это путем откладывания яиц, или путем деления, или почкования, или прорастания из семени, или как-то иначе. Наиболее показательна удельная рождаемость, определяемая как количество особей, появившихся в единице времени на одну особь в популяции (в демографии расчет ведется на одну женщину репродуктивного возраста)  . Реальная рождаемость в значительной степени зависит от факторов среды, поэтому она всегда меньше максимальной рождаемости, под которой понимают теоретический максимум рождаемости, определяемый только физиологией особей при оптимальных значениях всех факторов среды.
. Реальная рождаемость в значительной степени зависит от факторов среды, поэтому она всегда меньше максимальной рождаемости, под которой понимают теоретический максимум рождаемости, определяемый только физиологией особей при оптимальных значениях всех факторов среды.
Смертность DNm характеризует гибель особей в популяции и выражается числом особей, погибших в единицу времени. Удельная смертность характеризует уровень смертности для определенной группы особей, например, количество смертей за месяц на тысячу особей популяции  . Смертность также зависит от факторов среды и бывает обычно значительно выше минимальной смертности при идеальных условиях среды, которая определяется физиологией данного вида организмов - даже в идеальных условиях особи будут умирать от старости.
. Смертность также зависит от факторов среды и бывает обычно значительно выше минимальной смертности при идеальных условиях среды, которая определяется физиологией данного вида организмов - даже в идеальных условиях особи будут умирать от старости.
1.5.1.2. Кривые выживаемости
При оценке динамики используют еще один параметр - выживаемость. Если смертность в популяции общей численностью N особей обозначить через М, то выживаемость определится как разность N-M. Большой интерес представляют кривые выживания особей популяции в зависимости от их возраста.
Различают три общих типа кривых выживания (рис. 1.13). Выпуклая кривая 1 характеризует относительно низкую смертность на протяжении почти всего жизненного цикла популяции. К концу жизненного цикла смертность значительно возрастает. Такая кривая характерна для популяций, в которых основной акцент при размножении делается не на количество новорожденных особей, а на повышенную заботу о молодом поколении. Такого рода кривая характерна для многих высших животных, а также и для человека.

Рис. 1.13. Типы кривых выживания
Вогнутая кривая 2 характеризует повышенную смертность на ранних стадиях жизненного цикла, которая снижается, когда особи достигают определенного возраста. Такие кривые типичны для низших организмов и растений. Например, смертность среди молодых дубов и прорастающих желудей особенно велика, но когда дубы достигают определенных размеров, их смертность резко снижается. Аналогичные кривые характерны для рыб, где смертность среди мальков обычно гораздо выше, чем смертность среди взрослых особей. Повышенная смертность в раннем возрасте в таких случаях компенсируется повышенным количеством семян, икринок, яиц и т.п.
Промежуточные типы кривой выживания колеблются обычно вокруг средней линии 3. Одним из примеров может служить ступенчатая кривая 4, характерная для видов, у которых смертность значительно меняется на разных стадиях жизненного цикла. Примером могут служить некоторые виды насекомых, жизненный цикл которых проходит последовательно через несколько стадий: яйцо, личинка, куколка, взрослое насекомое.
1.5.1.3. Кривые роста популяции
Однако наибольшее значение для экологии имеют не сами показатели плотности или численности популяции, или ее возрастная структура, а скорости, с которыми меняются все эти показатели.
Если среда не лимитирует популяцию, то динамика численности популяции описывается дифференциальным уравнением: 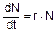 (скорость роста численности пропорциональна числу особей), которое имеет решение в виде: N=N0ert. Здесь N0 - численность в начальный момент времени, t - время, r - коэффициент роста популяции, определяемый как разность между удельной рождаемостью b (в расчете на одну особь) и удельной смертностью d в данный момент времени, то есть r=b-d. Максимально возможное значение коэффициента роста популяции rmax, достигаемое при наиболее благоприятных условиях среды, называется биотическим потенциалом популяции. Обычно биотический потенциал тем выше, чем ниже уровень организации организмов. Так дрожжевые клетки, размножаясь делением, при условии реализации биотического потенциала способны освоить весь земной шар за несколько часов. Крупным организмам с более низким потенциалом для этого потребовалось бы несколько десятилетий или даже столетий.
(скорость роста численности пропорциональна числу особей), которое имеет решение в виде: N=N0ert. Здесь N0 - численность в начальный момент времени, t - время, r - коэффициент роста популяции, определяемый как разность между удельной рождаемостью b (в расчете на одну особь) и удельной смертностью d в данный момент времени, то есть r=b-d. Максимально возможное значение коэффициента роста популяции rmax, достигаемое при наиболее благоприятных условиях среды, называется биотическим потенциалом популяции. Обычно биотический потенциал тем выше, чем ниже уровень организации организмов. Так дрожжевые клетки, размножаясь делением, при условии реализации биотического потенциала способны освоить весь земной шар за несколько часов. Крупным организмам с более низким потенциалом для этого потребовалось бы несколько десятилетий или даже столетий.
В соответствии с решением уравнения динамики численности популяции можно построить так называемую кривую роста популяции. Из уравнения видно, что это экспонента, поэтому такую кривую называют экспоненциальной (рис. 1.14).
Особенность экспоненциальной динамики в том, что рано или поздно относительно свободный рост численности популяции прекращается. Связано это, как правило, с исчерпанием какого-либо ресурса, на базе которого развивается данная популяция, например пространство или пища, вода. Причиной также могут служить внезапные заморозки или какой-то другой фактор среды, прекращающий сезон относительного благополучия. После этого численность популяции может катастрофически понизиться (пунктирная линия на рис. 1.14). Иногда вслед за подобным падением идет повторный рост численности, в результате формируется так называемая триггерная кривая роста, свойственная, например, однолетним растениям, некоторым насекомым, популяциям леммингов в тундре и т.п.
 |  |
Рис. 1.14. Типы кривых роста популяции
Подобные тенденции вызывают немалые опасения в плане будущего человеческой цивилизации. Несмотря на все прогнозы демографов, динамика численности людей не стабилизируется, оставаясь очень близкой к экспоненциальной кривой. Мы уже близки к разрушению собственной среды обитания, а значит, вполне возможно, что в достаточно скором будущем (возможно, при жизни нынешних поколений) нас может ожидать катастрофическое снижение численности населения Земли.
Обычно лимитирующее давление среды играет в динамике популяции роль отрицательной обратной связи, поэтому в уравнении динамики популяции появляется дополнительный сомножитель или слагаемое: 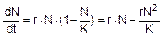 , где К – емкость среды – максимально возможная численность популяции при данных условиях среды, это верхняя асимптота, соответствующая верхнему пределу численности популяции после того, как в системе наступит равновесие (ее величина определяется средой). В данном случае скорость роста равна максимальной скорости роста (rN), определяемой биотическим потенциалом, умноженной на степень реализации максимальной скорости (1-N/K), которая тем ниже, чем больше плотность, или численность N, популяции. Поэтому такую динамику называют зависимой от плотности (численности), в отличие от независимой от плотности экспоненциальной динамики.
, где К – емкость среды – максимально возможная численность популяции при данных условиях среды, это верхняя асимптота, соответствующая верхнему пределу численности популяции после того, как в системе наступит равновесие (ее величина определяется средой). В данном случае скорость роста равна максимальной скорости роста (rN), определяемой биотическим потенциалом, умноженной на степень реализации максимальной скорости (1-N/K), которая тем ниже, чем больше плотность, или численность N, популяции. Поэтому такую динамику называют зависимой от плотности (численности), в отличие от независимой от плотности экспоненциальной динамики.
Решение этого дифференциального уравнения имеет вид:  , где a - постоянная интегрирования, определяющая положение кривой относительно начала координат. Кривая роста популяции, описываемая данным уравнением, называется логистической (рис. 1.14).
, где a - постоянная интегрирования, определяющая положение кривой относительно начала координат. Кривая роста популяции, описываемая данным уравнением, называется логистической (рис. 1.14).
Почти все математические модели роста популяций сильно уязвимы. В наиболее частом случае динамика популяции не идет ни по одной из этих кривых, реальная кривая роста располагается где-то в промежутке между этими кривыми. По инерции рост численности еще некоторое время может продолжаться даже после выхода за пределы верхней асимптоты, но затем после нескольких колебаний численность популяции, как правило, либо стабилизируется, либо совершает незначительные колебания относительно значения N=K. Иногда эти колебания могут быть достаточно существенными, их называют популяционными волнами или волнами жизни. Причины популяционных волн могут быть как внешними (периодические изменения факторов среды), так и внутренними, связанными особенностями межвидовых и внутривидовых отношений.
1.5.1.4. Экологические стратегии выживания
Стратегии – это стремление организмов к выживанию. Все многообразие стратегий заключено между двумя типами эволюционного отбора, которые обозначаются константами логистического уравнения: r- стратегия и К-стратегия.
Тип r-стратегия (r-отбор) – направлен прежде всего на повышение скорости роста популяции за счет: высокой плодовитости, ранней половозрелости, короткого жизненного цикла, способности быстро распространяться на новые места обитания и пережить неблагоприятное время в покоящейся стадии.
Тип К-стратегия (К-отбор) – направлена на повышение выживаемости в условиях уже стабилизировавшейся численности. Это отбор на конкурентоспособность, повышение защищенности от хищников и паразитов, повышение вероятности выживаемости каждого потомка, на развитие более совершенных внутривидовых механизмов регуляции численности.
Очевидно, что каждый организм испытывает на себе комбинацию r - и К – отбора, но r- отбор преобладает на ранней стадии развития популяции, а к-отбор характерен для стабилизированных систем. Но все-таки оставляемые отбором особи должны обладать достаточно высокой плодовитостью и достаточно развитой способностью выжить при наличии конкретных условий среды.
1.5.2. Территориальная структура популяций
Распределение особей в популяции может быть случайным, равномерным и групповым. Случайное распределение встречается в тех случаях, когда на популяцию воздействует одновременно множество достаточно слабых факторов среды. Необходимым условием для этого должно быть отсутствие тенденции к образованию групп (стайный инстинкт, семейные группы или нечто подобное). В случае же группового распределения сами группы обычно распределяются в пространстве случайным образом. Равномерное распределение отдельных особей или их групп (точнее близкое к равномерному) характерно в случаях с достаточно жесткой внутривидовой конкуренцией.
Примером случайного распределения может служить расположение личинок мучного хрущака в мешке с мукой, одиночные паразиты или хищники, например пауки, также распределяются обычно случайным образом. Случайное распределение в природе встречается относительно редко. Так например, распределение высоких деревьев в лесу уже нельзя назвать случайным. Обычно кроны таких деревьев смыкаются и конкуренция в борьбе за свет способствует выравниванию распределения.
У животных тенденция к равномерному распределению связана с явлением территориальности. Активность особей, пар или семейных групп многих животных обычно бывает ограничена определенным пространством, которое называют индивидуальным или семейным участком. Если эти участки активно охраняется, так что они практически не пересекаются друг с другом, то они называются территориями. Территориальность распределения наиболее ярко выражена у позвоночных.
Явление территориальности - это один из механизмов поддержания определенного динамического равновесия внутри популяции. Эволюция форм этого равновесия в конечном итоге приводит к идее так называемых “безопасных поселений”.
Безопасные поселения характеризуются большим скоплением особей на каком-то достаточно благоприятном для жизни участка. Это может быть остров посреди океана, целиком или частично занятый колонией птиц (птичьи базары); муравейники, термитники, пчелиные ульи, человеческие города - все это примеры безопасных поселений. Агрегация на центральном участке обеспечивает колонию своими преимуществами: повышенная защищенность, благоприятные условия для размножения, экономия затрат энергии на поддержание жизнедеятельности вследствие специализации. В то же время весь приток необходимых для жизни средств идет со стороны. Добыча пищи идет вовсе не на центральном участке, а на обширном пространстве, окружающем этот участок.
Отрицательным моментом подобных поселений является загрязнение центрального участка своими отходами, вытаптывание растительности, уплотнение почвы и т.п., что особенно ярко наблюдается в птичьих базарах. Слишком большая плотность популяции приводит к разного рода стрессовым явлениям. Слишком большая численность популяции требует расширения региона, за счет которого существует данное поселение.
Практически все сказанное про безопасные поселения целиком можно отнести и к нашим городам, разрастание которых грозит экологическими проблемами, недостатком ресурсов, поскольку уменьшает свободное пространство за городскими пределами, за счет которого собственно города и существуют.
ª Вопросы для самопроверки
1. Дайте определение понятию "популяция".
2. Какие количественные характеристики применяют для оценки динамики популяций ?
3. Что такое кривые выживаемости и роста популяции ?
4. Опишите территориальную структуру популяций.
5. Какие виды распределения особей в популяции вы можете назвать ?
6. Что такое сопротивление (давление) среды ?
7. Почему численность популяций из года в год остается примерно постоянной ?
8. Что такое давление жизни с точки зрения экологии ?
1.5.3. Особенности динамики численности человечества
Численность населения в настоящее время близка к 6,11 млрд человек (2001 г.). В природе для относительно крупных млекопитающих (к которым может быть отнесен человек) такая численность беспрецедентна. Численности видов животных в природе ограничиваются емкостью среды. Как правило, виды мелких животных имеют большую численность по сравнению с видами крупных животных. Для видов млекопитающих существует достаточно выраженная корреляция между массой тела и численностью особей. Рангу массы тела от 10 до 100 кг, к которому относится человек, соответствует максимальный размах значений видовой численности в пределах 107- 104. Современная численность видов ближайших родственников человека — человекообразных обезьян — близка к последней цифре. Вероятно, нормальная биологически обусловленная численность вида, к которому мы принадлежим, близка к 500 тысячам особей. В настоящее время она превышена на четыре порядка — в 10 тысяч раз.
На протяжение многих тысячелетий народонаселение мира росло чрезвычайно медленно. Считается, что к началу IV-го тысячелетия до н.э. оно составляло около 100 млн, к 1000 году н.э. едва достигало 300 млн, а к 1500 году увеличилось до 425 млн. Такие темпы соответствуют годовому приросту максимум до 0,7 %, т.е. до семи человек на 10000 за год. Начиная с эпохи Великих географических открытий темпы роста народонаселения заметно возросли и приблизились к экспоненциальной закономерности. Но затем нарастание числа людей на Земле, прослеженное уже на основании достаточно достоверных исторических данных, приобрело еще более необычный характер, стало гиперболическим. В период с 1600 по 1990 гг. рост населения Земли довольно точно (с максимальным отклонением в 5 %) описывается эмпирическим уравнением гиперболы:

 ,
,
где Y - год н.э.
По этому уравнению получается, что в 2030 г. численность должна стать бесконечной. Поскольку это невозможно, закон, по которому росло население в указанный период, должен в ближайшее время измениться. Действительно, за последние годы, с конца 80-х годов, относительный прирост почти перестал увеличиваться; рост численности приблизился к линейному со средним значением абсолютного прироста около 86 млн человек за год. С учетом линейного роста ожидается ежегодный прирост населения на 77 млн человек по крайней мере в течение следующего десятилетия. По прогнозам экспертов Фонда народонаселения ООН (1995 г.), этот темп сохранится до 2015 г., когда общая численность людей достигнет 7,5 млрд, а в 2050 г. численность населения Земли составит от 7,9 до 10,9 млрд человек, причем наиболее вероятна численность в 9,3 млрд человек.
Для оценки темпов роста населения используют следующие показатели:
1) ОКР, ОКС – общий коэффициент рождаемости/смертности, среднее число рождений и смертей на 1000 человек в год.
Общий означает, что данные цифры не учитывают, какую часть населения страны составляют пожилые и молодые люди, мужчины и женщины;
2) темпы прироста (убыли)  , %;
, %;
3) время удвоения T2 – период, необходимый для удвоения численности при современных темпах прироста
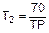 ;
;
4) количество населения через t лет можно рассчитать, зная ТР, по следующей формуле
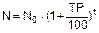
или с учетом экспоненциальной зависимости роста (как для популяций живых организмов)
 , где r=b-d – скорость роста, связанная с темпом роста как r=TP/100;
, где r=b-d – скорость роста, связанная с темпом роста как r=TP/100;
5) СКР – суммарный коэффициент рождаемости (или КФ – коэффициент фертильности) – среднее число детей, которые рождаются у женщины в течение жизни, или среднее число детей, приходящихся на одну семью.
Графическое представление о возрастной структуре популяции дает половозрастная пирамида – гистограмма для отображения полового и возрастного состава населения, демонстрирующая количество людей в каждой возрастной группе, обычно с разницей в 5 лет (рис. 1.15).
 |
Рис. 1.15. Пирамиды населения в странах с низким (прогрессивная) и высоким ВНП (стационарная) на душу населения, 1995 и 2025 гг.
(Для России характерна регрессивная половозрастная пирамида, т.к. темпы роста населения отрицательные. Молодые возрастные группы имеют меньшую численность, чем группы более старшего возраста)
Очень быстрое увеличение численности человечества в XX в. называют демографическим взрывом. Наприрост за последние 100 лет приходится три четверти современного числа людей на Земле. С экологической точки зрения рост человечества в XX в. имеет характер, больше свойственный г-стратегам — в основном примитивным организмам, у которых часто вслед за резкой вспышкой численности (демографическим взрывом) следует массовая гибель. Для отдельных популяций крупных К-стратегов (приматов, копытных, хищников), но никогда для вида в целом, максимальный размах численности иногда достигает 500 %.
Кроме особых факторов эволюции и истории человечества, значительное ускорение роста происходило в основном из-за снижения смертности от эпидемий и голода. В XX в. к этим факторам добавилось резкое снижение детской смертности в развивающихся странах, связанное с улучшением гигиенических условий и успехами медицины. Демографический взрыв обусловлен тем, что, начиная с середины столетия, снижение смертности значительно опередило снижение рождаемости во многих странах мира, причем наиболее резко — в развивающихся странах.
В 1990—1995 гг. общий коэффициент рождаемости в мире снизился до 24,6 %, общий коэффициент смертности составлял 9,8 %, коэффициент естественного прироста - 14,8 %. Примерно такие же параметры воспроизводства населения сохраняются и в настоящее. время. Это означает, что в среднем каждую минуту на Земле появляются 270 младенцев, умирают 110 человек разного возраста, население мира увеличивается на 160 человек. Каждые сутки в тесном орбитальном космическом корабле под названием планета Земля число пассажиров возрастает на 230 тысяч.
Темпы роста населения зависят от уровня развития страны. Численность населения высокоразвитых стран приближается к постоянному уровню, поэтому основной прирост населения в развивающихся странах. По абсолютной численности самый большой прирост в азиатских странах – Китай, Индия, Индонезия, наибольшая скорость прироста в Африке, Латинской Америке. Прирост составлял 4 % в год. Стабилизация численности будет наблюдаться при простом воспроизводстве: СКР=2,03 для развитых стран и СКР=2,20 для развивающихся стран (т.к. там выше детская смертность). В последние годы этот показатель равен для развитых стран 1,9, для развивающихся 4,1. Для России в 1995 г. СКР=1,4, ТР= -0,1 % в год.
Рост численности населения непосредственно влияет на реальное благосостояние (РБ), которое оценивается как
РБ = РЭ – РН,
где РЭ - экономический рост, РН - рост населения.
Например, РБ = 2 % - 0,6 % =1,4 % - для развитых стран, РБ = 1,6 % - 4 % = - 2,4 % - для развивающихся.
Некоторые пути решения проблем народонаселения:
1) увеличение производства продуктов питания,
2) более равномерное распределение сельскохозяйственной продукции по разным странам и группам населения,
3) медицинская помощь развивающимся странам,
4) повышение уровня образования женщин,
5) снижение рождаемости и государственное регулирование уровня рождаемости и др.
В большинстве развитых стран характерно в настоящее время развитие демографического перехода к стабилизации численности населения. Демографический переход сходен с фазой динамики численности, когда она асимптотически приближается к уровню емкости среды, и r -стратегия переходит в K-стратегию.
Выделяют 4 фазы перехода (рис. 1.16):
1) от примитивной стабильности (высокие смертность и рождаемость)переходят к состоянию, когда смертность и рождаемость быстро снижаются, но падение коэффициента смертности (ОКС) опережает падение коэффициента рождаемости (ОКР), поэтому прирост населения продолжает нарастать, достигая максимума – этот процесс называют демографическим взрывом;
2) смертность минимальна, рождаемость продолжает падать, прирост населения замедляется (фаза характерна в настоящее время для развивающихся стран);
3) повышение ОКС в связи со старением населения, замедление падения рождаемости. К концу 3-й фазы коэффициент рождаемости близок к уровню простого воспроизводства, а коэффициент смертности ниже этого уровня (характерно для развитых стран);
4) цивилизованная стабильность - коэффициент смертности повышается, сближается с коэффициентом рождаемости

Рис. 1.16. Одна из вероятных моделей четырех стадий демографического перехода к стабилизации численности
Это состояние соответствует уровню воспроизводства, при котором ОКР=ОКС=12, т.е. прироста населения не наблюдается, численность стабильна.
В составе такой популяции будет 23 % детей и юношей до 18 лет, 49 % людей трудоспособного возраста (18-60 лет) и 28 % людей в возрасте от 60 до 84 лет. На каждую 1000 человек будет 96 семей с возрастом родителей от 20 до 36 лет, в каждой из которых будет в среднем по 2 ребенка.
Одно популярное американское издание опубликовало следующие расчеты. Если бы все население Земли “сжать” до размеров деревушки с населением в 100 человек, а все существующие соотношения современного человечества остались бы прежними, то получилась бы следующая картина:
- в ней проживало бы 57 азиатов, 21 европеец, 14 представителей Северной, Центральной и Южной Америки, 8 африканцев,
- 70 из 100 были бы “цветными” (не белыми),
- 50 % всех богатств оказались бы в руках 6 человек, и все они были бы гражданами США,
- 70 человек не умели бы читать,
- 50 человек страдали бы от недоедания,
- 80 человек жили бы в жилищах, для проживания не приспособленных,
- только 1 человек имел бы университетское образование.
Прогноз: 90 % населения планеты со временем будет жить в развивающихся странах и, главным образом, в бедности.
ªВопросы для самопроверки
1. Какое влияние оказывает на биосферу рост народонаселения ?
2. Как определить темпы прироста населения и время его удвоения ?
3. Какие количественные показатели используют для определения темпов роста народонаселения ?
4. Каковы пути решения проблем роста народонаселения ?
&?Вопросы для самостоятельного изучения
1. Почему демографический взрыв (быстрый, экспоненциальный рост населения) не может долго продолжаться ?
2. Какие главные факторы накладывают ограничение на рост численности человечества или населения отдельных стран ?
3. Прокомментируйте с точки зрения экологии утверждение Т. Мальтуса о том, что человечество размножается в геометрической прогрессии, а производство продуктов питания увеличивается в арифметической прогрессии. В чем он был прав и в чем ошибался с точки зрения современной экологии ?