Обменная диффузия при посоле
1. Перераспределение соли и воды
В момент соприкосновения рассола с поверхностью продукта между ним и рассолом возникает обменная диффузия, которая приводит к перераспределению соли, воды и растворимых составных частей продукта между ними.
Этот процесс является наиболее важным, так как от количества соли зависит вкус и устойчивость продукта к действию микроорганизмов. Количество воды в продукте определяет его выход и консистенцию, а также концентрацию соли в тканевой жидкости, следовательно, и устойчивость к воздействию микроорганизмов. Переход растворимых составных частей продукта в рассол имеет отрицательное значение, так как продукт теряет некоторое количество ценных веществ.
Проникновение соли в ткань и перераспределение между тканью и рассолом происходит по меньшей мере двумя путями: а) осмотически через мембраны и перепонки, покрывающие внешнюю поверхность обрабатываемого участка ткани, б) через систему макро- и микрокапилляров, пронизывающих ткань во всех направлениях с последующим перераспределением соли и воды между этой системой и клеточными элементами ткани. При этом проникновение соли вторым путем происходит в первую очередь и с большей скоростью.
Кусок мышечной ткани при центрифугировании (2000 об/мин)отделяет ничтожно малое количество тканевой жидкости. Этот же кусок после посола при центрифугировании отделяет 10—15% рассола, заполняющего наиболее крупные поры ткани. Концентрация соли в таком рассоле в любой момент посола выше концентрации соли в тканевой жидкости, не отделяемой центрифугированием. Так, через 15 суток посола в 12%-ном рассоле концентрация соли в ткани составила 4,2%, а в центрифугате 5,7%; в 24%-ном рассоле эти концентрации соответственно оказались равными 10,9 и 11,3%.
Расчет, произведенный на основании результатов этих опытов, показал, что примерно на 10-е сутки посола количество воды, содержащейся в капиллярной системе, оказалось максимальным и составило: для 12%-ного рассола 12%, для 18%-ного рассола 14% и для 24%-ного рассола 18,5% к начальному содержанию влаги в мясе.
Основное количество воды, содержащейся в центрифугате, составляет перешедшая в капиллярную систему из клеток. За вычетом этого количества воды содержание влаги собственно в ткани (т. е. в клетках) на десятые сутки посола составило (в % к начальному содержанию влаги в мясе): для 12%-ного рассола — 88,2%, 18%-ного рассола — 83,6%, для 24%-ного рассола — 80,4%.
Следовательно, примерно на десятые сутки посола клеточная система максимально обезвоживается за счет перехода влаги из клеток в капиллярную систему, хотя общее содержание влаги в мясе оказывается больше начального (лишь в концентрированном рассоле оно несколько меньше).
После десяти суток посола количество воды, находящейся в системе макрокапилляров, начинает уменьшаться. Вместе с этим общее содержание воды в куске возрастает. Это свидетельствует о том, что клеточные элементы ткани начинают поглощать воду окружающего рассола и того, который удерживается в макрокапиллярах. Чем длительнее посол, тем меньше рассола остается в системе кровеносных и лимфатических сосудов. Это сказывается на величине потерь и на консистенции продукта при варке его и должно учитываться при выборе продолжительности посола мясопродуктов.
Систему макро- и микрокапилляров используют для ускорения посола тем или иным способом: вводят рассол в кровеносную систему через крупные сосуды, нагнетают его в систему капилляров, чередуя вакуум и давление, и т. д.
Наряду с перераспределением воды между структурными элементами тканей происходит ее перераспределение по формам связи с материалом: с течением времени посола возрастает доля влаги более прочно связанной и уменьшается доля связанной менее прочно. При посоле парного мяса количество прочно связанной влаги несколько больше, чем при посоле охлажденного. Увеличение доли прочно связанной влаги способствует повышению выходов и улучшению консистенции после варки, так как продукт лучше удерживает влагу.
Несмотря на сложность и многообразие описанных явлений, процесс посола может быть выражен уравнениями, сходными с теми, которыми пользуются для описания диффузионных процессов.
На рис. 46 представлен график изменения концентрации соли в рассоле и в погруженном в него куске мышечной ткани с течением времени. Концентрация соли в ткани выражена в % к весу воды и соли в ткани. Начальная концентрация хлорида в ткани — 0,2%. Судя по графику, концентрация соли в ткани растет, а в рассоле падает, приближаясь к какой-то конечной величине. (На графике выражена линиями АВ и СD). Эту величину можно получить путем расчета как гипотетическую равновесную концентрацию при очень большой продолжительности посола.
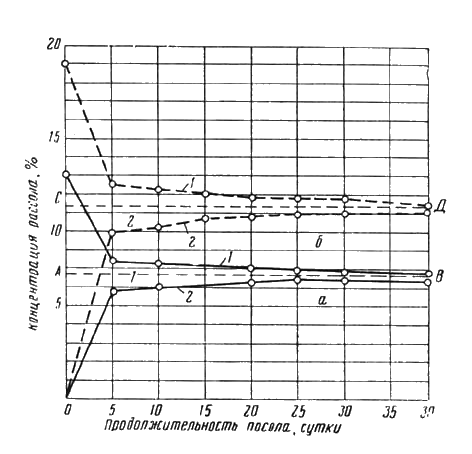
Рис. 46. Диаграмма изменения концентраций соли во времени:
а — для начальной концентрации рассола 12%; б — для начальной концентрации рассола 18%; 1 — в рассоле; 2 — в мясе.
Она является условной концентрацией суммарного количества соли в суммарном количестве рассола системы
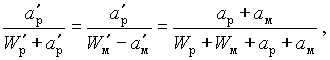 (V-1)
(V-1)
где
аp — количество соли в рассоле до посола;
ам — количество соли в мясе до посола;
Wр — количество воды в рассоле до посола;
Wм — количество воды в мясе до посола.
Те же символы с индексом соответствуют состоянию после посола.
Теоретически равновесное состояние, которое могло бы быть достигнуто по истечении бесконечно большого промежутка времени, должно характеризоваться правилом Доннана, согласно которому в присутствии заряженных частиц по одну сторону мембраны, не способных диффундировать через нее, по другую сторону должен обнаруживаться некоторый избыток ионов. Практически посол заканчивают задолго до наступления такого равновесия, когда в продукте еще сохраняется какой-то градиент концентрации. Так, например, при законченном посоле окороков, производимым простым погружением в рассол, обнаруживается следующее распределение концентраций соли в равных по толщине слоях в направлении от поверхности мышечной ткани к слою шпика
| Слой окорока | шпик | ||||
| Содержание соли, % | 6,0 | 5,0 | 3,0 | 2,5 | 1,0 |
Из графика (рис. 46) следует еще один практический вывод: движущей силой обменной диффузии между рассолом и животными тканями является разность концентраций соли. Это дает основание полагать, что несмотря на сложность хода посола в деталях, кинетика этого процесса в общем виде может быть описана уравнениями, принятыми для описания диффузионных процессов.
Акад. С. М. Липатовым показана правомерность использования уравнения Фика для описания кинетики диффузионных процессов в гелях. Согласно этому для нестационарного процесса, протекаюшего в трех измерениях, скорость диффузии
 (V-2)
(V-2)
где
с — концентрация:
t — время;
D — коэффициент диффузии;
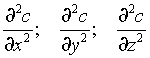 — градиенты концентрации в направлениях, перпендикулярных каждой из трех плоскостей сечения тела.
— градиенты концентрации в направлениях, перпендикулярных каждой из трех плоскостей сечения тела.
Для удобства рассмотрения хода процесса посола и его зависимости от различных технологических факторов можно исходить из представления о существовании лишь одного из трех направлений диффузии, т. е.
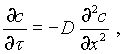 (V-3)
(V-3)
Такой подход имеет и практический смысл, так как в наиболее типичных случаях посола тело либо имеет форму пластины с площадью, во много раз превышающей толщину (шкуры, кишки), либо оно значительно более доступно диффузии с одной из сторон (окорок, грудинка и другие продукты, защищенные слоем жира и шкурой).
С. М. Липатовым дано решение дифференциального уравнения (V-3) для случая диффузии в гелях
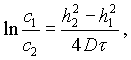 (V-4)
(V-4)
где
h1 и h2 — расстояния от поверхности тела, между которыми протекает диффузия;
c1 и c2 — концентрации на расстояниях h1 и h2 от поверхности тела.
При условии, если ход диффузии рассматривается на отрезке между поверхностью тела и точкой на глубине h, т. е. если h1 = 0, уравнение (V-4) примет вид
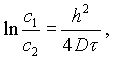 (V-5)
(V-5)
Как установлено экспериментально, распределение концентраций соли в мясе при мокром посоле для любого времени (до 39 суток) удовлетворяет этому уравнению.
При мокром посоле продуктов процессы перераспределения соли и воды складываются из трех одновременно протекающих фаз:
1) перераспределения соли и воды между рассолом и продуктом,
2) перераспределения соли и воды в рассоле,
3) перераспределения соли и воды внутри продукта.
Соль из рассола в продукт и внутрь его переходит при всех условиях диффузионно-осмотическим путем. Перенос соли в толще рассола путем диффузии может происходить лишь при условии состояния полного покоя для всей системы рассол — продукт. При таких условиях устанавливаются различные градиенты концентрации: в слое рассола, между рассолом и продуктом и внутри продукта, как это изображено кривой 1 на рис. 47. При этом концентрация соли вблизи поверхности продукта оказывается значительно ниже средней для всего объема рассола. В силу этого соль из рассола в продукт переходит с наименьшей скоростью, когда отсутствует конвекция.
Если рассол перемешивается искусственно вследствие конвекции, перекладки продукта или по другим причинам, диффузионный перенос соли в рассоле вытесняется молекулярным (конвективным) переносом. При интенсивности перемешивания, достаточной для быстрого выравнивания концентрации соли в рассоле, практически устанавливается концентрация, близкая к средней. В этом случае диффузионный перенос соли в рассоле будет иметь место только в пределах пограничного слоя, толщина которого зависит от скорости движения рассола. Различные градиенты концентрации устанавливаются: в пограничном слое, между пограничным слоем и поверхностным слоем продукта и внутри продукта, как это изображено кривой 2 на рис. 47. При этом перенос соли из рассола в продукт и внутри продукта при прочих одинаковых условиях осуществляется с максимальной скоростью, зависящей от интенсивности перемешивания.
В связи с тем, что состояние покоя системы рассол — продукт постоянно нарушается, анализ диффузионно-осмотической сущности собственно посола без большой погрешности можно производить, отправляясь от концентрации рассола непосредственно вблизи пограничного слоя.
На основании уравнения (V-5), изменение концентраций соли, вызываемое диффузионным переносом в пограничном слое, может быть выражено уравнением
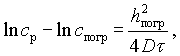 (V-6)
(V-6)
где
ср — концентрация рассола у пограничного слоя;
спогр — концентрация соли в пограничном слое у поверхности тела;
hпогр — толщина пограничного слоя;
D — коэффициент диффузии соли в растворе (при 4° D = 0,710 см2 в сутки).
Изменение концентрации соли в результате диффузионно-осмотического переноса из пограничного слоя в поверхностный слой тела может быть выражено уравнением
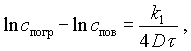 (V-7)
(V-7)
где
спов — концентрация соли в поверхностном слое продукта;
k1 — коэффициент пропорциональности, учитывающий изменение скорости диффузии при переходе из пограничного слоя в продукт вследствие изменения свойств среды.
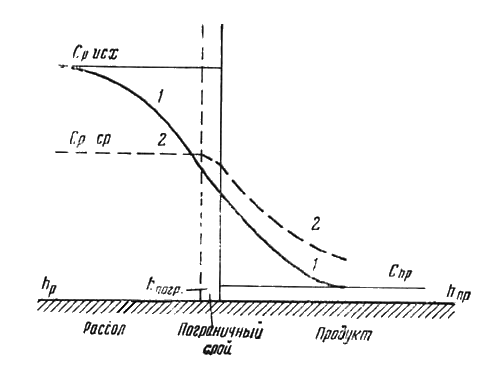
Рис. 47. Диаграмма изменения концентрации соли в рассоле, пограничном слое и в толще продукта:
1 — для состояния покоя; 2 — при перемешивании; Срисх — исходная концентрация рассола; Спр — продукта; Срср — средняя концентрация рассола, достигаемая перемешиванием; hp — толщина слоя рассола; hпогр — толщина пограничного слоя; hпр — толщина слоя продукта.
Изменение концентрации соли, вызываемое диффузионным переносом внутри продукта, может быть выражено уравнением
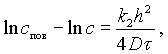 (V-8)
(V-8)
где
с — концентрация соли на глубине h;
k2 — коэффициент пропорциональности, учитывающий падение скорости диффузии внутри продукта.
Изменение скорости движения частиц соли в толще продукта в сравнении со скоростью движения в чистом растворе обусловлено по крайней мере следующими причинами:
а) движущиеся частицы преодолевают множество полупроницаемых перегородок,
б) движение частиц происходит в коллоидных системах, а не в чистой воде и тормозится силами молекулярного сцепления,
в) вода, в которой происходит диффузия, занимает лишь часть площади сечения продукта, следовательно, поверхность диффузии составляет лишь часть геометрической поверхности.
Суммируя уравнения (V-6, V-7, V-8) и переходя от натуральных логарифмов к десятичным, получим
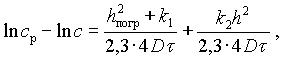 (V-9)
(V-9)
Физический смысл величины k2 может быть понят из уравнения (V-8), написанного в виде
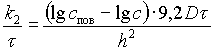 .
.
Из него следует, что эта величина характеризует уменьшение градиента концентрации, приходящиеся на единицу пути диффузии в продукте и вызываемое ее сопротивлением движению частиц. Величина k2 выражает свойства продукта, которые могут влиять на скорость диффузии соли внутри его. Это подтверждается тем, что k2 меньше для диффузии в направлении вдоль мышечных волокон, чем для диффузии в направлении поперек волокон; для размороженного мяса с частично разрушенной структурой тканей, чем для охлажденного; для мышечной ткани, чем для жировой. Она зависит также от концентрации рассола, влияющей на плотность тканей: чем она выше, тем больше величина k2. Поэтому, строго говоря, ее нельзя считать постоянной величиной, поскольку концентрация рассола меняется в ходе посола. Однако для технических целей можно руководствоваться средней величиной k2 применительно к какой-либо начальной концентрации рассола.
По аналогию величина (hпогр+k1) характеризует сопротивление движению частиц в пограничном слое рассола и на переходе из него в поверхностный слой продукта. Следовательно, она является, с одной стороны, функцией состояния рассола (покой, движение), с другой — свойств поверхностного слоя продукта. Это подтверждается тем, что указанная величина зависит как от скорости движения рассола, так и от тех же факторов, которые влияют на величину k2.
В практике толщина пограничного слоя может колебаться в широких пределах. В состоянии полного покоя для системы рассол — продукт по существу весь слой рассола приобретает свойства пограничного слоя. При интенсивном перемешивании его толщина уменьшается до незначительной величины. В общем, искусственная циркуляция рассола увеличивает скорость посола в 1,1—1,5 раза в зависимости от интенсивности циркуляции. При периодическом перемешивании или при перекладывании продукта его толщина — величина переменная. Влияние состояния рассола и условий посола на толщину поверхностного слоя не изучалось. В связи с этим величину (hпогр + k1) расчленить пока невозможно. Принимая ее равной средней величине для обычных условий влажного посола мяса и выразив ее через k1, уравнение (V-9) можно представить в виде
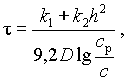 (V-10)
(V-10)
Это уравнение и может быть рекомендовано для различных случаев посола в пределах технических потребностей. Его справедливость иллюстрируется графиком, приведенным на рис. 48, на котором зависимости, выражаемые уравнением, в координатах “h2” и “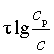 ” дают прямую.
” дают прямую.
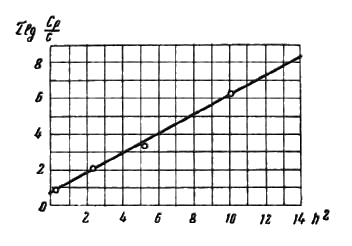
Рис. 48. Графическое изображение зависимостей между концентрациями соли, временем посола и толщиной слоя мяса в координатах “h2” и “ ”
Примерные величины k1 и k2 для свиного охлажденного мяса приведены в табл. 80 [17].
Таблица 80
| Условия посола | Начальная концентрация рассола, % | k1 | k2 |
| Рассол проникает вдоль волокон | 1,6 | 3,0 | |
| 2,0 | 3,2 | ||
| 2,6 | 4,0 | ||
| Рассол проникает поперек волокон | 2,6 | 3,2 | |
| 2,9 | 3,7 | ||
| 3,3 | 4,5 |
При диффузии вдоль от мышечных волокон средняя скорость проникновения соли в мышечную ткань в обычных условиях посола составляет около 0,059 см/ч, при диффузии поперек от волокон — около 0,039 см/ч.
Увеличение коэффициентов сопротивления k1 и k2 с возрастанием начальной концентрации рассола объясняется уменьшением доли воды в том сечении, через которое протекает диффузия.
Это уменьшение вызывается большей степенью обезвоживания тканей в более концентрированных рассолах.
Влияние замораживания и размораживания мяса на величину коэффициентов сопротивления зависит от степени разрушения тканей. В опытах по посолу размороженного мяса были получены следующие результаты:
| Температура замораживания, °С | —10 | —17 | —24 |
| Уменьшение коэффициентов сопротивления относительно охлажденного мяса (в %) до |
Скорость распределения соли между рассолом и тканью, а также и внутри ее зависит от температуры. В пределах от 0°С до 50° С эта зависимость определяется только влиянием температуры на коэффициент диффузии соли D:скорость посола возрастает прямо пропорционально коэффициенту диффузии. Влияние температуры на коэффициент диффузии может быть учтено с помощью уравнения [15, 16]:
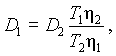 (V-11)
(V-11)
где
Т1 и Т2 — абсолютная температура;
h1 и h2 — коэффициенты вязкости рассола, соответствующие указанным температурам;
D1 и D2 — коэффициенты диффузии соли для тех же температур.
Увеличение скорости посола с повышением температуры можно характеризовать следующими цифрами (принимая скорость посола при 4° С равной единице) [16]:
| Температура, °С | Кратность увеличения скорости |
| 1,7 | |
| 2,1 | |
| 2,7 | |
| 3,4 |
В течение первых шести-семи суток посола скорость диффузии соли заметно меняется от жесткости воды, используемой для приготовления рассола (жесткая вода уменьшает скорость диффузии). Затем влияние жесткости уменьшается.
Одновременно с перераспределением соли между рассолом и продуктом начинается и перераспределение воды, характер и продолжительность которого мало изучены. При консервировании посолом количество воды, остающееся в продукте, сказывается на его устойчивости к гнилостной микрофлоре; при выработке соленых пищевых продуктов влияет на выход, вкус и структурно-механические характеристики изделий.
Характер перераспределения воды в процессе посола с учетом его двух фаз существенным образом зависит от времени и концентрации рассола, а также интенсивности просаливания и размеров образца продукта.
При любой концентрации рассола вначале посола происходит обезвоживание продукта, вызываемое более высоким осмотическим давлением рассола в сравнении с осмотическим давлением тканевой жидкости. Степень обезвоживания тем больше, чем выше концентрация рассола. Она достигает максимума при обычных условиях посола для мясопродуктов приблизительно на пятые-седьмые сутки, для шкур — к 18—48 ч в зависимости от их веса. После этого начинается обводнение продукта, обусловленное нарастанием в нем концентрации соли. Обводнение продолжается в течение всего времени посола, хотя и при понижающейся интенсивности. Максимальное количество воды поглощается мясом, когда содержание соли в нем достигает 4—5% к его весу. Равновесное состояние наступает в сроки, лежащие далеко за пределами тех, которые установлены принятой технологией посола.
В начальный период посола интенсивность перехода воды из продукта в рассол превышает интенсивность перехода соли из рассола в продукт. В связи с этим при повышенной концентрации рассола вначале вес продукта уменьшается. В последующем, когда начинается его обводнение, вес начинает возрастать. Если используют слабые рассолы, вследствие относительно небольшой степени обезвоживания в начальный период вес увеличивается почти с самого начала посола.
Осмотическое давление насыщенного рассола превышает давление набухания животных тканей. Поэтому в тех случаях, когда в течение всего периода посола концентрация рассола поддерживается на уровне насыщения, продукт обезвоживается все время вплоть до наступления равновесного состояния. Так обстоит дело, например, при посоле кишок в насыщенном рассоле. Это может быть использовано для устранения подсолки шкур, необходимой при их тузлуковании в ненасыщенных рассолах [8].
Практически одновременно с перераспределением воды между рассолом и продуктом начинается перераспределение воды между системой капилляров (кровеносной и лимфатической системой) и межклеточным пространством с одной стороны и морфологическими структурными элементами (клетками и волокнами) с другой. При этом, как уже было показано, в начальный период посола происходит обезвоживание морфологических структурных элементов.
При посоле мяса обезвоживание клеток продолжается в течение некоторого времени и после того как начинается обводнение всего продукта в целом и достигает максимума примерно на 10 сутки. Практическая значимость разрыва во времени между максимумом обезвоживания продукта и максимумом обезвоживания морфологических структурных элементов состоит в том, что, несмотря на возрастание веса сырого продукта, весовые потери после варки могут оказаться наибольшими.
Количество воды, удерживаемое соленым мясом, зависит от его водосвязывающей способности, которая, в данном случае связана и с концентрацией рассола. При значениях рН, близких к изоэлектрической точке белков мяса, она возрастает с повышением концентрации рассола до 10%, а затем уменьшается. Влияние рН на водосвязывающую способность мяса существенное при низких концентрациях рассола, мало заметно при высоких. При значениях рН 5,5 и ниже потери воды при посоле и варке наибольшие.
Степень и скорость обезвоживания продукта в рассоле зависит от величины осмотического давления рассола, которое в свою очередь определяется ионной концентрацией электролитов вне зависимости от их природы. В некоторых случаях, например при консервировании шкур, осмотическое давление даже концентрированного рассола не обеспечивает желательной скорости и степени обезвоживания продукта. Для таких процессов целесообразнее пользоваться смешанными рассолами, содержащими два или более электролита, например концентрированным раствором поваренной соли с добавлением какого-либо другого электролита, значительно повышающего осмотическое давление рассола.
Уравнения (V-9) и (V-10) пригодны также и для характеристики динамики изменения концентрации соли при посоле свиного шпика в рассоле. Однако интенсивность ее изменения в шпике в несколько раз меньше, чем в мышечной ткани. В отличие от посола мышечной ткани в шпике постоянно увеличивается количество воды, связываемой его белковой частью. Поглощение воды шпиком происходит до тех пор, пока его влажность не достигнет определенной величины, зависящей главным образом от содержания белковых веществ и незначительно от концентрации рассола.
Одновременно с водой из рассола в жировую ткань диффундирует и поваренная соль, количество которой на единицу веса воды, поглощаемой ею, в начале посола несколько больше, чем в последующем. Это соотношение зависит от количества и первоначальной концентрации рассола.
При посоле мясопродуктов сухой солью вначале образуется рассол на поверхности за счет влаги самих продуктов. С этого момента между продуктом и рассолом возникает обменная диффузия, аналогичная той, которая имеет место при мокром посоле. Поэтому для скорости посола имеет большое значение количество и время возникновения рассола на поверхности. В свою очередь они зависят от влажности, структуры и других свойств поверхности продукта, обрабатываемого сухой солью. Быстрее всего образуется рассол на поверхности мышечной ткани, медленнее на жировой и наиболее медленно на внешней поверхности шкуры.
Если на поверхности продукта в течение всего времени посола имеется избыток соли, концентрация рассола на ней близка к концентрации насыщенного раствора. Осмотическое давление такого раствора способно преодолеть давление набухания белков мяса. Поэтому при сухом посоле продукт всегда теряет воду в количестве, которое зависит от избытка соли. Так, при посоле шкуры с избытком сухой соли, содержание влаги в ней уменьшалось следующим образом:
| Продолжительность посола, ч | ||||
| Содержание влаги, % | 50,4 | 48,7 | 40,3 | 36,8 |
Если продукт длительное время находится в соприкосновении с сухой солью, он практически теряет всю осмотическую и капиллярную влагу, а воды остается не более 70% к весу сухого остатка. Эта величина лежит в пределах адсорбционной влажности большинства животных белков. Животные ткани после длительной обработки сухой солью, перенесенные в рассол, набухают хуже.
2. Особенности обменной диффузии в живых клетках
При посоле действию повышенного осмотического давления подвержены также и живые микробные клетки. В отличие от мертвых животных тканей они в рассоле ведут себя, как открытые термодинамические системы: равновесное состояние с окружающей средой в клетках наступает в процессе обмена веществ и поддерживается непрерывной затратой энергии, которая освобождается в результате окислительных процессов в клетках. Вследствие этого живые клетки не успевают достигнуть состояния равновесия Доннана с окружающей средой. Между ними и средой сохраняется более или менее значительная разница осмотических давлений, величина которой зависит от интенсивности обмена веществ в клетке. Клетки, способные к большим энергетическим затратам, могут также поддерживать большую разность осмотических давлений, т. е. сохранять сравнительно низкое осмотическое давление в протоплазме при наличии высокого осмотического давления в окружающей среде. Они выживают в концентрированных солевых растворах.
Клетки, не способные к интенсивному обмену веществ в условиях повышенного осмотического давления, не способны также и к поддержанию большой разности осмотических давлений между протоплазмой и средой. Они погибают вследствие резкого повышения осмотического давления в протоплазме.
Вторая особенность поведения живых биологических систем в растворах электролитов заключается в избирательном характере диффузии ионов в клетки. В таких системах равновесное соотношение концентраций веществ по обе стороны мембраны (клеточной оболочки) разное для различных ионов. Оно зависит не только от их подвижности, но и характера участия в химических и физико-химических процессах, протекающих внутри клетки. Вследствие этого, хотя протоплазма и проницаема для всех ионов, в клетке происходит предпочтительное накопление одних ионов в сравнении с другими.
Для нормальной жизнедеятельности клеток в растворе электролита необходима быстрая нейтрализация образующихся в протоплазме водородных ионов путем обмена с катионами извне. Если в растворе имеются ионы калия, в клетках возникает диффузионный потенциал, способствующий обмену ионов водорода на ионы калия и затрудняющий их обмен на ионы натрия. Клетки пекарских дрожжей в растворе калиевой соли оставались живыми, а рН раствора KCl вследствие выхода водородных ионов снижался до 3,0 и менее. Замена калия ионами натрия прекращала нормальную жизнедеятельность клеток и вызывала их гибель. При этом снижение рН раствора было значительно меньшим. Аналогичное явление наблюдалось и в опытах с одной из культур молочнокислых бактерий.
Различие в отношении биологических систем к разным ионам в данном случае обусловлено различием в сольватации ионов калия и натрия. Более мощная сольватная оболочка иона натрия уменьшает притягательное действие противоионов в протоплазме, проявляющееся только на определенных расстояниях. В результате ионы натрия оказываются в менее благоприятных условиях в смысле их хемосорбции протоплазмой, нежели ионы калия.
Живая мышца, погруженная в раствор хлористого натрия, вначале обменивала лишь часть ионов калия на эквивалентное количество ионов натрия и сохраняла жизнеспособность при 2—3° С в течение четырех-пяти суток. После этого происходило извержение всех ионов калия с заменой их ионами натрия, сопровождавшееся значительным набуханием и гибелью мышцы.
Таким образом, поведение живых биологических систем, в том числе макро- и микроорганизмов, в растворах электролитов обусловливается не только величиной осмотического давления раствора, но также природой ионов и биологическими особенностями организмов. Оно находится в зависимости от продолжительности воздействия электролита и находит свое выражение либо в гибели организмов, либо в их приспособлении к новым условиям.
3. Перераспределение растворимых составных частей продукта
В процессе посола в рассол переходят белковые, экстрактивные и минеральные вещества, способные растворяться в рассоле. Количество веществ, переходящих в рассол различно, и зависит от условий посола (крепости и количества рассола, его продолжительности и т.д.). В табл. 82 приведены данные, характеризующие потери наиболее важных составных частей мышечной ткани при мокром посоле говядины.
В рассоле обнаружены как альбумины, так и глобулины. Для потерь белковых веществ существенное значение имеет концентрация рассола: потери повышаются с ее увеличением до 10— 12%, а затем уменьшаются. При концентрации рассола 25% они минимальны.
Таблица 82
| Крепость посола, % | Продолжительность посола, сутки | Потери к первоначальному содержанию в мясе, % | ||
| белковых веществ | азотистых экстрактивных веществ | фосфорных соединений, % (Р2О5,) | ||
| 2,2 | 17,9 | 17,2 | ||
| 2,3 | 25,7 | 18,2 | ||
| 4,9 | 14,7 | 17,0 | ||
| 1,8 | 16,7 | 16,0 | ||
| 1,9 | 24,8 | 20,4 |
Потери относительного количества азотистых веществ (главным образом белков) зависят от категории мяса, и чем она выше, тем меньше эти потери (табл. 83).
Таблица 83
| Мясо | Потери белковых веществ через 20 суток, г/кг мякоти | |
| смешанный посол | сухой посол | |
| II категории | 5,9 | 4,5 |
| I категории | 5,6—6,2 | 2,8—4,6 |
Так как белковые вещества не способны диффундировать через стенки клеток, их потери вызваны только переходом в рассол белков, заполняющих кровеносную систему, и белков разрушенных клеток. В связи с этим величина потерь в известной мере зависит от полноты обескровливания мяса и степени разрушения тканей.
Белки соединительной ткани — коллаген и эластин — в рассол не переходят. Коллагеновые волокна лишь сильно набухают; набухание достигает максимума примерно на 20-е сутки посола.
Потери других (в том числе экстрактивных) веществ не зависят от концентрации рассола и достигают максимума в момент установления равновесия между мясом и рассолом (см. табл. 82).
В числе экстрактивных веществ находятся также и некоторые водорастворимые витамины: теряется до 35% фолиевой кислоты, 15 —20 витамина В1 и незначительное количество витамина В2.
Удаление растворимых белковых веществ при посоле шкур и кишок играет положительную роль, так как способствует устойчивости консервирования и улучшению качества.