Вторичная структура ДНК
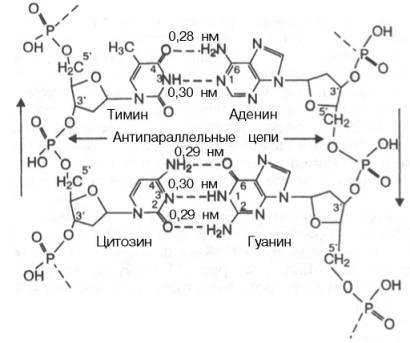
Исходя из данных рентгеноструктурного анализа и опираясь на правила Чартгоффа в 1953 году британские биохимики Уотсон и Крик предложили модель строения ДНК. Согласно этой модели ДНК построена из из двух полинуклеотидных цепей, антипарралельно ориентированных друг около друга ( одна цепь идет в направлении 5¢ -- 3¢ , другая 3¢ –5¢ ) и на всем протяжении связанных водородными связями. Каждый нуклеотид принимает участие в образовании водородных связей. Эти связи образуются за счет специфического взаимодействия между аденином и тимином, гуанином и цитозином. Причем Аденин и тимин гуанин и цитозин находятся в разных цепочках. Между аденином и тимином образуется 2 связи, между гуанином и цитозином – 3.
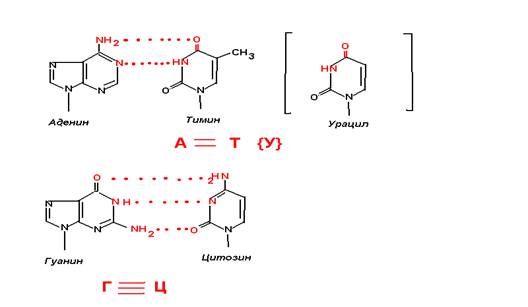 |
Экспериментальные данные показывают, что на разрыв водородных связей в молекуле ДНК, обогащенной гуанином и цитозином, требуется большие затраты энергии, чем ДНК богатой аденином и тимином.
Первичная структура одной цепи комплементарна первичной структуре другой цепи. Если в положении n одной цепи находится адениловая кислота, то в комплементарном участке другой цепи находится только тимиловая кислота и ни какая другая. Т.О. зная первичную структуры одной цепочки всегда можно установить первичную структуру другой.
Обе цепи закручены в спираль, имеющую общую ось. Каждый шаг такой спирали равен 3.4 нм. и на него приходится примерно 10 нуклеотидных пар.
Длина водородных связей между основаниями составляет около 0,3 нм. Таким образом, комплементарными оказываются не только отдельные основания, но и дезоксирибонуклеотидные цепи ДНК в целом, способствующие образованию весьма компактной структуры и стабилизации всей молекулы. диаметр биспиральной молекулы равен 1,8 нм.
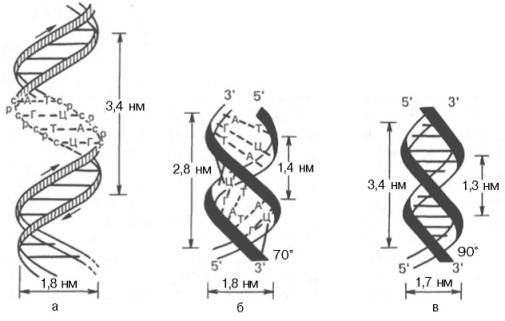
Необходимо указать, что конфигурация двойной спирали ДНК сильно меняется в зависимости от количественного содержания воды и ионной силы окружающей среды. Методами рентгеноструктурного анализа доказано существование по крайней мере 6 форм ДНК, названных А-, В-, С-, D-, Е- и Z-формами. Конфигурация двух из них в простейшей форме представлена на рис. 3.1, б и в. Можно увидеть, что у А-формы наблюдается некоторое смещение пар оснований от оси молекулы к периферии, что отражается на размерах (2,8 нм – длина одного витка, в котором вместо 10 содержится 11 мононуклеотидов; меняется расстояние между нуклеотидами и др. В настоящее время есть основание считать, что между А- и В-формами ДНК осуществляются взаимные переходы при изменении концентрации соли и степени гидратации. В-форма ДНК больше всего подходит к модели Уотсона и Крика. Предполагают, что в А-форме ДНК выполняет роль матрицы в процессе транскрипции (синтез РНК на молекуле ДНК), а в В-форме – роль матрицы в процессе репликации (синтез ДНК на молекуле ДНК).