Теории происхождения фаун
Прежние и современные очертания континентов и океанов, несомненно, оказали огромное влияние на современное фаунистическое и флористическое расчленение Земли. При изучении геологического и палеонтологического прошлого нашей планеты выдвигалось несколько точек зрения с обоснованием современного распространения растений и животных.
Первые воззрения, объясняющие распространение организмов на планете, строились на утверждении, что основа очертаний материков всегда оставалась неизменной или была близкой к современной конфигурации.
Одной из первых теорий, объясняющих географическое распределение организмов, была теория мостов суши, имевшая много сторонников преимущественно в XIX в. Согласно этой теории, сходство флор и фаун различных континентов объясняется тем, что в разное время эти регионы были соединены континентальными мостами. Так, близость фауны и флоры Индостана, о. Шри-Ланка, Мадагаскара и Африки объяснялась существованием материка Лемурии, соединявшего эти территории. Родственные связи флоры Западной Африки и Южной Америки объяснялись существованием материка Южной Атлантиды. Некоторая общность животного и растительного мира Европы и Северной Америки — существованием Северной Атлантиды, сходство биоты Северной Америки и Восточной Азии — существованием материка Берингии и т. д. Однако с современных геологических позиций только последний пример получил подтверждение. Теория мостов суши была опровергнута на основании исследования строения дна океанов, поэтому от этой теории в ее общем виде отказались, несмотря на кажущуюся простоту объяснения сходства флоры и фауны.
На смену ей пришла теория фиксизма (постоянства континентов). Согласно этой теории, очертания материков и океанов менялись лишь в своих второстепенных чертах: наступление океана на сушу и суши на океан наблюдалось только в пределах континентальной ступени (в пределах подъема и падения уровня Мирового океана). Основа же очертаний суши всегда оставалась неизменной. При опускании уровня океана обнажалось дно, позволяющее организмам расселяться на новые территории. Однако и эта теория не в состоянии объяснить многих разрывов ареалов, которые могли возникнуть только в результате заметных изменений соотношения между океанами и континентами.
Теория дрейфа материков разработана немецким ученым А. Вегенером в начале XIX в. Суть ее в том, что материковые глыбы из кремния и алюминия плавают в более глубоком слое Земли, состоящем преимущественно из кремния и магния, частично погружаясь в него. Все материки в палеозое составляли единую сушу — Пангею.
Под влиянием вращения Земли Пангея начала разламываться на части, которые стали смещаться в направлении к западу и югу. Еще в триасе Пангея (200 — 160 млн лет назад) существовала, но уже в юрское время (160 — 90 млн лет назад) возникли линии разлома, и связи между материками начали прерываться. Связи между Индией и Австралией существовали до юрского периода, между Африкой и Южной Америкой, Африкой и Индией — до эоцена (10 млн лет назад), между Австралией и Южной Америкой, Австралией и Антарктидой — примерно до конца палеогена (6 — 7 млн лет назад), между Европой и Северной Америкой, Антарктидой и Южной Америкой — до четвертичного периода (1 млн лет назад).
А. Вегенер видел подтверждение этой теории даже в сходстве очертаний противоположных берегов океанов, например, американского и африканско-европейского берегов Атлантического океана. Для доказательства существования этого горизонтального дрейфа он направился в глубь Гренландии, где и погиб.
Эта теория хорошо объясняла большинство известных биогеографических фактов и особенностей распространения животных и растений: миграции угрей на икрометание из рек Европы и Северной Америки в один район (Саргассово море), наличие представителей американского семейства кактусовых в лесах Западной Африки. Родство флор и фаун южной части Африки (Капская область), южной части Южной Америки, Австралии и островов Антарктиды легко объясняются положением этой теории относительно постепенного расхождения материков. Но эта теория не соответствует современным представлениям о строении Земли, поэтому была забыта на долгие годы. Однако ни одна из пришедших ей на смену теорий не смогла дать обоснование известным биогеографическим фактам.
Существовали также теория пендуляций (качаний), согласно которой земной шар качается вокруг своей оси, что вызывает перемещения материков, и теория расширения Земли, которое якобы вызвало растрескивание Пангеи.
Геологами в XX в. была выдвинута эквивалентная теории движения материков А. Вегенера теория тектоники плит (рис.6), или новая глобальная теория, основанная на системе срединно-океанических хребтов с их осевыми рифтовыми долинами (рис.7). Оказалось, что эти хребты и остальное океаническое ложе сравнительно молоды: они сформированы всего за последние 150 — 200 млн лет, в то время как геологическая история Земли насчитывает 3500 — 4000 млн лет.
Возникла гипотеза, что земные оболочки разделяются на хрупкую и жесткую литосферы, полностью включающую земную кору и часть верхней мантии до глубины примерно 70— 100 км; глубже нее простирается так называемая астеносфера,вещество которой находится в состоянии, близком к плавлению, и способно перетекать. Литосфера на большей части Земли непрерывна. Она разорвана на части лишь в узких местах, соответствующих сейсмически активным районам земного шара: срединным океаническим хребтам, островным дугам и молодым горным цепям. В этих поясах лежат области разлома плит. В срединноокеанических хребтах сквозь разрывы литосферы астеносфера, изливаясь, вызывает «растекание морского дна» (растекающиеся по периферии плиты).
Таким образом, в отличие от теории А. Вегенера, при которой предполагалось, что континенты плавают в магме, в настоящее время полагают, что земная кора вместе с верхней частью мантии представляет единое целое, предвигающееся по астеносфере. В астеносфере развиваются компенсационные конвекционные течения. Расхождение плит вызывает либо их уничтожение в одной зоне, либо столкновение в другой.
Молодые горно-складчатые сооружения типа Гималаев возникли при столкновении плит. Большинство геологов насчитывает 6 крупных тектонических плит: евразиатская, американская, африканская, австралийская, антарктическая (все континентальные) и тихоокеанская (океаническая). Иногда выделяются также более мелкие плиты — Карибская, Филиппинская и др. Толщина плит — 70—100 км, скорость их движения по различным подсчетам от 1 до 10— 16 см в год. Таким образом, Океаны имеют неодинаковый возраст. Наряду с древними океанами — Тихим и Тетисом (последний отделял южный, тогда еще единый материк — Гондвану, в состав которого входили современная Антарктида, Австралия, Южная Америка, Африка и Индостан, — от северного материка Лавразии), имеются и молодые — Атлантический и Индийский.
Эта теория основана на всей совокупности наук о Земле и объясняет большую часть закономерностей распространения организмов.
2. Главные этапы развития животного мира. Понятие о фауне. Видовой состав. Экологическая природа видов. 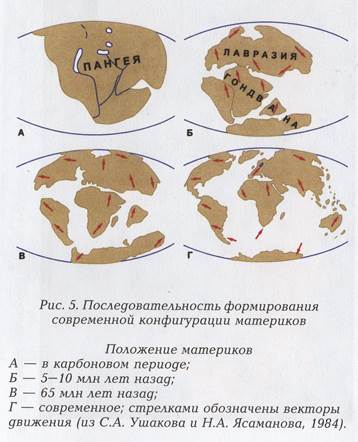
Современное распространение животных и растений является результатом общего палеогеографического развития земного шара, начиная с палеозоя (570 млн лет назад) и до наших дней. Главные этапы этого развития можно коротко охарактеризовать следующим образом (рис. 5):
1. Начиная с палеозоя и до триасового, или юрского, периода мезозоя Земля имела вид одного (Пангея) или двух (Лавразия и Гондвана) блоков. Группы животных, эволюционировавших к тому времени, могли свободно расселяться по южным материкам, включая Мадагаскар, Индию и Новую Зеландию. Такое же расселение было между Европой и Северной Америкой (около 200— 160 млн лет).
2. После раскола Гондваны и удаления друг от друга ее фрагментов началось независимое развитие их фаун. Фрагменты Гондваны еще до конца мезозоя были соединены, в частности, тропики Южной Америки с Африкой, южные части Южной Америки с Австралией и Новой Зеландией, а также с Антарктидой ( 160 — 40 млн лет).
3. В течение продолжительного третичного периода (40 — 5 млн лет) к огромному Евроазиатскому материку присоединился Индостан (фрагмент Гондваны). Кроме того, указанный материк постоянно сохранял связи с Северной Америкой в районе нынешнего Берингова пролива. Тропики южной Азии в третичном периоде были главным эволюционным центром доминирующих и наиболее конкурентоспособных групп животных, активно расселявшихся и вытеснявших на своем пути древнюю фауну. Так произошло с древней гондванской фауной Индии. Фауна североазиатского умеренного пояса, родственная фауне южных частей Азии, населила Европу и Северную Америку, образовав здесь эндемичные роды и виды. Юго-восток Северной Америки стал вторичным центром эволюции многих групп, вышедших из Азии. Позднее некоторые из них продвинулись в Южную Америку.
4. В миоцене произошло соединение Африки и Азии, что открыло путь в Африку многим обитателям тропических лесов и саванн — выходцам из Южной Азии. Будучи эволюционно молодыми и прогрессивными формами, они почти полностью вытеснили гондванскую фауну, и в конце третичного периода Африка стала главным центром развития и расселения многих групп животных. После образования Красного моря Африка снова отделилась.
5. Южная Америка, фрагмент Гондваны, к началу третичного периода (40 млн лет) ненадолго восстановила былое соединение с Северной Америкой, что открыло путь к расселению северных групп животных, особенно млекопитающих, на юг. Это соединение вскоре нарушилось, и Южная Америка в течение значительной части третичного периода оставалась изолированным континентом, вроде нынешней Австралии. В это время здесь развивались особые фауны, состоявшие из групп гондванского происхождения, а также выходцев из Северной Америки. В плиоцене (1,5 млн лет) снова произошло соединение Северной и Южной Америки, и новые волны переселенцев внедрились в южноамериканскую фауну. Однако они лишь частично вытеснили автохтонную фауну, так что она сохранилась гораздо полнее, чем фауна Африки.
6. Другой фрагмент Гондваны —Австралия — дрейфовал на северо-восток и в миоцене приблизился к Азии. Через цепочку островов произошло вселение на север Австралии таких азиатских животных, которые были способны пересечь морские проливы. Тем не менее нелетающие звери (за исключением семейства мышиных), большинство амфибий и все строго пресноводные рыбы на австралийский материк так и не попали.
7. В течение долгих веков географической изоляции континентальных блоков их фаунистическая изоляция не была полной, т. к. многие животные преодолевали разделяющие их пространства активным или пассивным путем. Это расселение облегчалось и тем, что материки не были удалены друг от друга на большие расстояния. Так, с эоцена до плиоцена изоляция Северной и Южной Америки была полной, и комплексы зверей, амфибий и строго пресноводных рыб развивались независимо друг от друга, но обмен между фаунами птиц был постоянным. Между южными континентами в области южной умеренной зоны и в настоящее время продолжается взаимопроникновение фаун, чему способствуют западные ветры и циркумантарктическое морское течение, несмотря на громадные расстояния. Конечно это касается только мелких животных, особенно членистоногих.
Фауна— эволюционно-исторически сложившаяся совокупность видов животных, обитающих в той или иной области (районе, местности). Так, фауна европейской части России на равных основаниях включает северного оленя, степного орла, щуку, бабочку-махаона, губку бодягу, в то время как места обитания их совершенно разные. Вместе с тем нельзя причислять к местной фауне виды, сознательно завезенные человеком и содержащиеся в зоопарках, аквариумах и т. д. Виды же, завезенные случайно, а также преднамеренно, но одичавшие или существующие без помощи человека, необходимо учитывать в составе местной фауны. К таковым, в первую очередь, относятся вредители, попавшие из других стран или районов (колорадский жук). Разумеется, при изучении истории фауны и путей ее формирования подобные виды приходится выделять в категорию привнесенных (случайно или преднамеренно).
Фауна характеризуется определенными, свойственными только ей признаками, позволяющими сравнивать ее с другими фаунами.
Видовой состав— самый важный признак любой фауны. Каждое фаунистическое исследование начинается с учета видов, обитающих в пределах изучаемого района, т. е. с инвентаризации фауны. Количество видов, входящих в состав фауны, отражает ее богатство. Изучение видового состава фауны сколько-нибудь обширного района требует длительного времени и коллективных усилий со стороны многих специалистов. К примеру, фаунистические исследования в Беларуси, начатые в XVIII в., продолжаются до настоящего времени.
Экологическая природа составляющих фауну видов. Например, для фауны тропического леса характерно наличие большого количества видов, приуроченных к деревьям. С одной стороны, это лазающие формы млекопитающих, птиц, рептилий и амфибий, многих насекомых и других животных. С другой — виды, связанные с деревьями трофическими отношениями, как то: плодоядные формы, потребители листвы, потребители и разрушители древесины. Большинству животных тропической фауны свойственна круглогодичная активность. Фауне степей присущи другие экологические типы — животные бегающие и роющие, проводящие зиму в состоянии спячки, питающиеся жесткой травой, семенами злаков и др.
Степень самобытности фаунывыражается в наличии эндемичных представителей. Чем выше систематический ранг эндемиков, тем самобытнее фауна. Так, присутствие в составе фаун эндемичных отрядов и семейств отражает длительность развития фауны в условиях изоляции. В то же время наличие только эндемичных видов свидетельствует об относительной молодости фауны. Примером древней и самобытной фауны служит фауна Австралийской области, в составе которой насчитывается 8 эндемичных семейств млекопитающих (сумчатые) и не менее 3 эндемичных семейств птиц, не считая большого количества эндемичных родов всех классов позвоночных животных. Самобытна и фауна Неотропической области. На территории Южной Америки обитают представители многих эндемичных семейств (17 из них млекопитающие, 26 — птицы), множество родов рептилий, амфибий, насекомых и т. д. Вместе с тем в пределах Голарктического царства, куда относится и фауна Беларуси, всего несколько эндемичных семейств позвоночных животных, хотя эндемичных родов немало. В целом голарктическая фауна значительно беднее и менее оригинальна, чем упомянутые выше.
3. Структура и сравнительный анализ фауны. Систематическая структура. Уровень эндемизма.
Каждая фауна познается через количественное соотношение различных систематических единиц, иначе говоря, обладает определенной систематической структурой — специфичным набором видов, родов, семейств и более высоких таксонов.
Для суждения о структуре фауны и сопоставления ее со структурой других фаун необходимо иметь большой объем данных по всем группам животных.
Для характеристики особенностей распространения элементов фауны отдельных территорий используют термины: эндемичные, реликтовые, автохтонные, бореальные, тропические, арктические, средиземноморские, среднеазиатские, нагорно-азиатские, европейско-сибирские и др. Термин «заносные» означает, что данные виды натурализовались благодаря заносу извне и не являются единой географической группой.
При сравнительном анализе фаун учитывают свойственные им признаки: количественное соотношение различных таксонов, уровень эндемизма ее представителей, экологические особенности распространения, а также происхождение.
Наличие или отсутствие конкретных таксонов и их количество в данной фауне можно наглядно отразить с помощью гистограмм, по процентному соотношению числа видов сравниваемых фаун, что позволяет выявить доминирующие таксоны.
Уровень эндемизма характеризует специфический компонент любой фауны и абсолютно отличает ее от других фаун. Если широко распространенные виды указывают на связи между фаунами, то эндемики отличают их друг от друга. Наиболее высок процент эндемизма в островных фаунах, на континентах — в районах с сильно расчлененным рельефом, т. е. в горных странах. По таксономическому уровню эндемизма можно судить о самобытности и древности фауны.
Обилие эндемиков отражает длительность развития фаун, поэтому в тех частях земного шара, где экологическая обстановка существенно не изменялась с третичного периода, сохраняется большое количество автохтонных эндемичных видов. Примером могут служить влажные экваториальные леса Южной Америки, Западной Африки и Юго-Восточной Азии. За время непрерывного развития этих фаун в них успели обособиться не только эндемичные виды, но и роды и даже виды семейства животных. Наоборот, молодые фауны, сформировавшиеся в областях, недавно освободившихся от четвертичного оледенения (север Европы или Северной Америки), практически не содержат сколько-нибудь значительного количества эндемиков.
Уровень эндемизма в разных фаунах различен. В фауне Австралии, например, 8 эндемичных семейств сумчатых млекопитающих и не менее 100 эндемичных родов птиц (из 270). В фауне неотропиков (Южная Америка) 17 эндемичных семейств млекопитающих (80% эндемичных видов) и 26 эндемичных семейств птиц (78% эндемичных родов). В то же время фауна всей внетропической Евразии имеет лишь по одному эндемичному семейству млекопитающих и птиц.
К реликтовым эндемикам (палеоэндемики) относятся животные, ареал которых в прошлом был гораздо больше, а в настоящее время они сохранились лишь на отдельной территории. Реликтовые эндемики занимают в фауне, как правило, изолированное положение. Реликтовые эндемичные роды часто включают лишь несколько или всего-навсего один вид (монотипичные роды). Экологическая обстановка их «устраивает» относительно. Конечно, если бы она полностью не отвечала их потребностям, они бы давно вымерли. Распространение этих видов в ареале носит пятнистый характер, они узко локальны и связаны с ограниченными биотопами. Наличие в фауне реликтовых эндемиков говорит о том, что благоприятная для них обстановка сохраняется длительное (в геологическом смысле) время, пусть даже на небольшом пространстве. Замечательным примером реликтового эндемизма служит гаттерия Spheodon punctatus — единственный представитель семейства клинозубых Sphenodontidae отряда клювоголовых Rhynchocephalia рептилий, живущий в Новой Зеландии.
Особенности распространения на рассматриваемой территории входящих в нее таксономических групп животных: горные, пустынные, околоводные, фоновые, редкие, космополиты и пр.
Чтобы узнать вероятное происхождение видов, пути попадания в состав фауны, требуется установить, какие элементы фауны возникли в пределах изучаемой территории, какие попали в результате расселения из других центров.
Если будет доказана автохтонность того или иного вида, следует выяснить, когда он возник, поскольку вопрос, откуда он проник, отпадает (он образовался на месте). Для решения данной проблемы нужны сведения об экологическом соответствии автохтонного вида современной обстановке и палеонтологические материалы.
У иммигрантов (аллохтонный вид) необходимо установить, откуда они проникли, когда вошли в состав фауны, какими путями совершалось их продвижение. Подобные вопросы требуют изучения ареалов и их динамики. Это позволит определить направление миграции вида. Обилие иммигрантов всегда указывает на молодость рассматриваемой фауны, обилие аборигенов — на ее древность.
Возраст фауны.В состав любой фауны входят элементы, различающиеся не только по географическому распространению, но и по возрасту. Часть из них сформировалась на месте, другие попали в фауну в результате расселения из первичных центров, но все они возникли в различные эпохи истории земного шара. В связи с этим важно установить:
1. Возраст вида или рода (время возникновения его как вида, рода и т. п.);
2. Время включения его в состав данной фауны.
Если вид относится к автохтонам, ответ на первый вопрос одновременно станет ответом и на второй. Для аллохтонных видов (мигрирующих) проблема решается по-другому. Вид может быть очень древним по происхождению, но в состав изучаемой фауны мог войти недавно, в процессе расселения из первичного очага. Определить абсолютный возраст его удается редко и только при наличии палеонтологического материала. Для большинства же животных этого материала нет.
Как правило, в фауне обычно выделяется несколько категорий видов в зависимости от степени соответствия их современным экологическим условиям, а значит, и от относительной давности пребывания их в составе данной фауны. Это прогрессивные, консервативные и реликтовые виды.
Для прогрессивных видов фауны современные условия среды благоприятны; численность их высока, а границы ареала имеют тенденцию к расширению (признаки биологического прогресса). К ним относятся прогрессивные эндемики, возникшие недавно в пределах территории, занятой фауной, и аллохтонные виды, недавно проникшие в данную область, и находящиеся в процессе расширения ареала. Это, например, заяц-русак в европейской части России, индийский скворец-майна в Средней Азии, канареечный вьюрок в Западной Европе и др.
К консервативной категории, как правило, относится преобладающее большинство видов любой фауны, находящихся в относительном равновесии с условиями существования. Ни признаков угасания, что выразилось бы в сокращении ареала, ни признаков экспансии (расширения ареала) консервативные элементы не проявляют. Численность их популяций колеблется около какой-то средней величины; они являются устойчивыми компонентами основных биоценозов и хорошо приспособлены к различным местам обитания. Несмотря на «стаж» пребывания в составе фауны, эти виды в настоящее время успешно развиваются.
Реликтами считаются виды (или роды), уже прошедшие эпоху расцвета и сократившие или сокращающие ареал. Современные условия существования не совсем соответствуют их экологическим требованиям — это основной фактор, определяющий выделение реликтовых форм. Процветание последних сегодня невозможно, о чем свидетельствует уменьшение численности их популяций и отступание от прежних границ ареалов.
Реликты могут быть и очень древними по своему абсолютному возрасту (гаттерия) и более молодыми (так называемые ледниковые реликты — заяц-беляк в Центральной Европе).
Генезис фауны.Каждая фауна имеет свою историю. Формирование фауны, или фауногенез, продолжается очень длительное время, измеряемое иногда целыми геологическими периодами. Лучшую опору для познания прежнего облика фауны дают ископаемые находки. Они и являются прямыми доказательствами пребывания какого-нибудь вида в составе изучаемой фауны в определенный период ее развития. Примерами могут служить находки скелетов мамонта в Сибири или окаменевших костей динозавров в Монголии.
Проанализировав способы зарождения и развития фаун на основе прямых и косвенных доказательств, Э. Майр (1965) предложил различать 5 типов фауногенеза:
Автохтонная адаптивная радиация — активный процесс видообразования на основе первоначально небольшого количества видов-основателей в географически изолированных районах. Таким путем возникла богатая фауна сумчатых млекопитающих Австралии, птиц и наземных моллюсков Гавайских островов и др. Вновь образующиеся таксоны заполняют свободные экологические ниши островов, архипелагов и даже изолированного материка — Австралии.
Повторяющаяся или непрерывная колонизация из одного источника — тип формирования фауны на островах, когда колонисты попадают с одного материка. Подобным образом произошло заселение вулканических Галапагосских островов только из Южной Америки. Подобный путь формирования фауны обычен и для некоторых озер, связанных единой речной сетью.
Повторяющуюся или непрерывную колонизацию из нескольких источников как форму фауногенеза следует считать наиболее обычной. Именно таким образом складывались фауны большинства регионов Земли. К примеру, в Африке различают автохтонные группы гондванского происхождения, а также группы, вышедшие из Южной Америки, южноазиатские и европейские группы животных. Время вселения этих групп в африканскую фауну, естественно, разное: группы южноамериканского происхождения могли попасть на африканский материк только в меловом периоде (130 млн. лет назад), а европейские — только в плейстоцене (2 — 2,5 млн лет назад). В островных фаунах иногда сочетаются формы, проникшие из разных, иногда прямо противоположных источников. Подобное можно наблюдать на островах Южной Атлантики (острова Св. Елены, Вознесения и др.), где в составе фауны есть вселенцы из Африки и Южной Америки.
Слияние двух фаун, территориально изолированных друг от друга эффективными барьерами, в одну (после исчезновения барьеров) сравнительно не часто встречающийся тип фауногенеза. К примеру, озеро Виктория (Центральная Африка) в промежутках между влажными периодами за счет частичного усыхания разделялось на многочисленные мелкие озера. Во время самостоятельного существования этих озер населявшие их фауны развивались независимо друг от друга, что и привело к появлению в каждом из них самостоятельных видов. При любом же новом увлажнении климата озера объединялись в одно. Следовательно, современная фауна Виктории является результатом слияния нескольких фаун.
Приспособление к специфическому местообитанию как особый тип фауногенеза. Вследствие колонизации из одного или из нескольких источников происходит естественный отбор с последующей вспышкой видообразования в специфических условиях (в океанических впадинах, глубоких пещерах, на изолированных горных вершинах и т. д.). Это приводитк образованию оригинальных фаун. Так, по всей вероятности, осуществлялся фауногенез в Австралии, где в меловом периоде произошло повторное вселение млекопитающих либо из Южной Америки (через Антарктиду), либо из Южной Азии. Переселенцами были примитивные сумчатые. Затем в миоцене или плиоцене (от 27 до 10 млн лет назад) из Азии в Австралию проникли плацентарные звери — водяные мыши. Сумчатые успешно прошли пугь адаптивной радиации, а водяные мыши слабо подвергались этому процессу. Итак, процесс фауногенеза можно выяснить по доле участия автохтонов и мигрантов в составе современных фаун.
Наличие большого количества широко распространенных видов при низком проценте эндемизма говорит в пользу преобладания миграционных процессов в фауногенезе.
И наоборот, количественное преобладание эндемиков, особенно высокого ранга, свидетельствует о доминировании местных длительных формообразовательных процессов.
Островные фауны
Во-первых, острова интересны из-за относительной простоты структуры их биоценозов и трофических связей между видами, что облегчает изучение сообщества; во-вторых, из-за необычных, порой удивительных форм жизни. Исследование островных фаун выявило способности наземных животных к преодолению морских рубежей, а также закономерности формирования новых фаун, изолированных от материковых.
Все разнообразие островов Мирового океана можно свести к нескольким основным типам. Главным критерием выделения типов является происхождение островов. Чаще всего они бывают океаническими и материковыми.
Океанические острова никогда не были частями материков, а образовались либо в результате поднятия дна океана, либо при извержениях подводных вулканов и застызания лавы, либо вследствие жизнедеятельности рифообразующих кораллов. Примерами подобных островов являются архипелаги Микронезии и Полинезии в Тихом океане, о. Св. Елены и др.
Материковые, или континентальные, острова представляют собой части материков, отделившиеся от них в определенную геологическую эпоху. К ним относятся Британские, Большие Зондские (Ява, Суматра, Калимантан и др.), Тайвань, Новая Гвинея, Тасмания и многие другие. Каким же образом формировалась фауна островов различного типа?
1. Остров материкового происхождения при отделении от материка сохраняет тот или иной (но не полный) набор местообитаний и связанных с ним видов животных и растений. Этим определяется зоогеографическое своеобразие таких островов: с момента их отделения от материка начинается дифференцировка некоторых видов, образуются местные формы (подвиды, виды, реже роды) в результате местного эволюционного процесса. С другой стороны, происходит обеднение первоначальной фауны за счет исчезновения ряда видов. Наконец, на материковых островах отсутствуют виды или роды, возникшие на материках или проникшие на них после отделения острова. Часть видов фауны и флоры эти острова получили от соответствующего материка, часть — за счет активного и пассивного расселения определенных животных. Островная фауна в любом случае будет беднее предковой материковой.
2. На острове океанического происхождения первое время после его образования вообще нет никаких организмов. Современный органический мир таких островов формируется за счет видов, попавших на них путем залетов или заносов, т. е. активным или пассивным путем. В некоторых же случаях в фауну входят обитатели океана, приспособившиеся к жизни на суше. К последним можно отнести некоторых рыб, раков (например, пальмовый вор) или пресноводных моллюсков. Следует подчеркнуть, что фауна океанических островов от начала своего образования и до достижения устойчивого состояния непрерывно обогащается. Признаки островных фаун. Независимо от происхождения фауна острова несет «островной» отпечаток, степень которого зависит от площади острова, расстояния его до ближайшей суши, от климатической зоны, в которой он располагается, и т. д. Иначе говоря, существуют конкретные признаки островных фаун— это бедность, выраженный эндемизм, ограниченное количество архаических форм и, наконец, адаптивная радиация в рамках небольшого количества эндемичных групп.
Относительная бедность (систематическая) островных фаун объясняется тем, что площадь острова всегда меньше площади ближайшего материка, и часть биотопов на нем отсутствует или занимает по сравнению с материком меньшую территорию. На острове может не быть крупных рек, а саванны или леса могут иметь меньшую протяженность. Отсутствие на острове каких-то групп или видов объясняется тем, что на острове отсутствуют характерные для них биотопы, либо они вымирают вследствие недостатка условий для поддержания необходимой численности популяций. К факторам, вызывающим островное обеднение, относится и антропогенный.
На океанических островах могли укорениться лишь виды, способные активным или пассивным путем пересекать водные преграды. Поскольку такой способностью обладают далеко не все животные, бедность фауны островов можно предсказать заранее. На океанических островах обитают летающие животные — птицы, летучие мыши, многие насекомые. Из наземных нелетающих форм раньше других здесь появляются грызуны, из рептилий — гекконы. И те, и другие могут заноситься на «плотах» из плавающих стволов деревьев, вырванных или сломанных тропическими ураганами и выброшенных в море.
На океанических островах отсутствуют строго пресноводные рыбы, многие семейства рептилий, по давляющее большинство амфибий и млекопитающих, прежде всего копытных, из беспозвоночных — скорпионы и ряд представителей других классов, которые ни активным, ни пассивным путем не могли расселиться через море. Это, в первую очередь, пресноводные моллюски (двустворчатые — беззубки, перловицы), отдельные брюхоногие, пресноводные раки. Живущие в пресных водах океанических островов рыбы и беспозвоночные являются потомками морских предков или относятся к родам и семействам, постоянно обитающим в пресной воде, но способным переносить засоление и пересекать моря.
Даже крупные древние континентальные острова имеют относительно бедную фауну. Так, на Мадагаскаре обитают только два семейства амфибий, но нет строго пресноводных рыб, сухопутных черепах и пресноводных, из змей отсутствуют аспидовые и гадюки и такие характерные для Африки семейства птиц, как птицы-секретари, цесарки, журавли, птицы-носороги, дятлы, вьюрковые и др. Из млекопитающих на Мадагаскаре нет и типично африканских семейств — слонов, носорогов, обезьян, кошек, гиен, антилоп, трубкозубов и т. д. Особенно следует подчеркнуть отсутствие копытных, кроме речной свиньи, и обилие хорошо летающих рукокрылых, родственных как африканским, так и индо-малайским формам.
Следовательно, бедность фауны — общий и характерный признак всех островов, проявляющийся тем в большей степени, чем остров меньше по площади и дальше находится от материка или другого острова.
Эндемизм — является результатом изоляции островов от других участков суши, поэтому большинство островов, кроме очень молодых по времени образования, отличаются уровнем видового или родового эндемизма. Самые древние острова (Мадагаскар, Новая Зеландия и др.) имеют даже эндемичные семейства. Степень эндемизма тем больше, чем древнее остров и чем он более удален от материка.
Таким образом, изоляция является причиной не только бедности фауны, но и ярко выраженного островного эндемизма. Так, на Мадагаскаре, отделенном о Африки морским проливом шириной 400 км и ставшем островом еще в мезозое, живут эндемичные семейств млекопитающих (тенреки из насекомоядных, три семейства лемуров) и птиц (нелетающие пастушковы куропатки, филапитты, а в прошлом голубь дронт и страус эпиорникс), многочисленные эндемичные роды и большое количество видов всех классов животныа Фауна птиц Новой Каледонии содержит более 20% эндемичных видов и одно эндемичное семейство.
Геологически молодые, расположенные близко о материка острова (например, Британские, Сахалин] эндемиков высокого ранга не имеют, хотя эндемичные подвиды или даже виды здесь обитают. Следователь но, процент эндемизма и систематический ранг эндемиков могут служить показателем возраста образования острова. Согласно зоогеографическим данным, Британия, и Сахалин стали островами только в четвертичном периоде, что подтверждается геологическим исследованиями.
Богатство фауны острова, степень ее эндемизма могут зависеть, кроме того, от его площади, разнообразия рельефа и других природных условий, а также от климатической зоны, в которой он находится. Все это создает большее или меньшее разнообразие экологических ниш. Так, низменные коралловые острова Тихого океана имеют малый набор экологических ниш и бедную фауну. Гористые же материковые острова от личаются большим разнообразием природных условш и, соответственно, богатой фауной. Экстремально хо лодный климат островов полярного бассейна сам по себе препятствует развитию богатой фауны независимо от их размеров и возраста.
Архаичность форм. Островные фауны нередко являются последним убежищем архаичных видов животных. Такие виды немногочисленны и встречаются на Мадагаскаре, Новой Зеландии, Сулавеси, а также на Антильских островах. В одной из оригинальнейших наземных фаун — новозеландской — вообще отсутствуют аборигенные млекопитающие, но имеются древнейшие виды: из рептилий — гаттерия, из амфибий —гладконогие лягушки лиопельмы (2–3 вида). В состав современной мадагаскарской фауны входят древние тенреки (30 видов), настоящие лемуры, индри и руконожки (из низших приматов), игуаны (из игуановых рептилий).
Отделение островов от материка или образование их другим способом происходило в разные периоды, в том числе и до возникновения на материках прогрессивных групп животных. Вот почему на островах сохранились древние обитатели, не имеющие врагов и конкурентов, которые перешли на положение оттесненных реликтов.
Кроме того, определенные виды животных, возникшие на островах, не могут жить на материке, даже если бы они туда и попали. Это в первую очередь нелетающие птицы. Они есть в Новой Зеландии, Новой Гвинее, на Мадагаскаре и прилегающих к нему малых островах, а также в Австралии, которую можно рассматривать как гигантский остров. Сохранение нелетающих птиц на островах возможно до тех пор, пока туда не будут завезены собаки или не начнется прямое преследование их человеком, что и произошло с гигантскими страусами моа в Новой Зеландии или нелетающими голубями на островах Маврикий и Реюньон вблизи Мадагаскара.
В фауне островов часто поражают ее необычные представители, с одной стороны — гиганты: варан на острове Комодо; исполинские, до нескольких центнеров, черепахи на острове Маврикий; истребленные человеком птицы эпиорникс на Мадагаскаре и дронты на Реюньоне (также достигали гигантских размеров); с другой стороны — карлики, такие как мелкая форма тигра на Зондских островах, мелкие барсуки и лисицы на острове Сардиния и т. д. (ископаемые слоны на острове Мальта достигали размеров кабана). Островная популяция лошадей, предки которых были завезены с материка, выродились в мелкую породу — пони. Причины как одного, так и другого явления до настоящего времени не выяснены.
Адаптивная радиация. Некоторые виды, попав на острова, настолько изменяются, что иногда уже мало чем напоминают предковую форму. Классическим примером адаптивной радиации являются гавайские птицы — цветочницы. Их предок, американский вьюрок, залетев на Гавайские острова, не нашел здесь конкурентов и дал начало нескольким эволюционным ветвям, нынешние представители которых по внешнему виду и образу жизни напоминают зябликов, медососов, пищух, дубоносов и дятлов. Все эти формы занимают определенные экологические ниши, что исключает возможность повторной адаптивной радиации в последующие эпохи, а также внедрение в гавайскую фауну настоящих дятлов или дубоносов.
С течением времени животное население островов существенно изменяется. Пополнение зависит, как уже отмечалось, от площади острова и расстояния от него до материка или другого источника заселения.