Фотоконверсия фитохрома. Фитохромы А и В
Фитохром представляет собой димер белка с ковалентно связанной хромофорной группировкой из четырех пиррольных колец (А, В, С и D). Молекула хромофора синтезируется сначала как замкнутый тетрапиррол (гем), а затем размыкается и встраивается в белок. Хромофор имеет структуру, близкую к фикобилинам (см. подразд. 3.3.3).
Как и ожидалось, хромофор фитохрома (фитохромобилин) существует в двух основных спектральных формах, которые переходят друг в друга. Форма Ф660 (красная) улавливает «красные кванты» света и переходит в Ф730 (дальнюю красную). Это объясняет эффект К/ДК-обратимости: при освещении красным светом форма Ф660 исчезает (форма Ф730 накапливается). Если после этого осветить растение светом с длиной волны 730 нм, форма Ф730 перейдет в Ф600. Последовательные вспышки приводят к тому, что сначала весь фитохром переходит в одну форму, а потом в другую и т. д. Для физиологической реакции важна именно последняя форма фитохрома, т. е. последняя короткая вспышка, которой освещали растение.
Главным событием, приводящим к фотоконверсии форм фитохрома, является поворот кольца D относительно кольца С (цис-транс-изомеризация в метиновом мостике) (рис. 7.28).
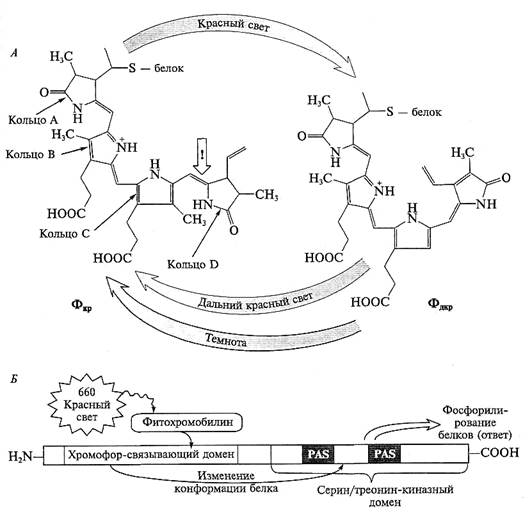
Рис. 7.28. Структура фитохрома:
А — структура хромофорной группировки и ее изменение при фотоконверсии форм фитохрома (см. также рис. 7.26);
Б — функциональные домены белковой части фитохрома
Эта реакция приводит к изменению конформации белкового окружения хромофора и димера в целом. Первичная форма возбужденной хромофорной группировки оказывается недостаточно стабильной и самопроизвольно через ряд этапов переходит в более стабильные формы, причем промежуточные формы для перехода из Ф660 в Ф730 отличаются от промежуточных форм для обратного перехода (см. рис. 7.26).
У арабидопсис известны 5 генов, отвечающих за биосинтез белкового компонента фитохрома. По количеству белка преобладают фитохром А и фитохром В, различающиеся не только белковой частью, но и спектром поглощения (эффект различного окружения хромофора). Фитохромы С, D и Е — минорные. В белковой части всех фитохромов ближе к N-концу есть консервативный домен, ковалентно связывающий фитохромобилин. При его повреждении хромофор не встраивается в фитохром. С-концевые участки отвечают за передачу сигнала (обычно есть два гастидинкиназных домена).
По сравнению с Ф(В) в спектре поглощения фитохрома А более широкие пики, у Ф(А)660 максимум поглощения — 660, однако широкое плечо выходит в дальнюю красную область. Это означает, что Ф(А)660 переходит в Ф(А)730 не только при облучении квантами с λ= 660 нм, но и при освещении дальним красным светом высокой интенсивности. Следовательно, у реакций, вызываемых Ф(А), нет К/ДК-обратимости. Фитохром А отвечает как за ответы на сверхнизкие интенсивности красного света (λ = 660 нм), так и за высокоэнергетический ответ на дальний красный (λ = 730 нм). В спектре поглощения фитохрома В пики более узкие, у формы Ф(В)660 плечо не заходит в дальнюю красную область. Фитохром В отвечает за ответы на низкую интенсивность и обеспечивает К/ДК-обратимость. Наличие фитохромов А и В, отличающихся спектрами поглощения, «позволяет» растению различать (1) полную темноту, (2) свет, обогащенный ДК-квантами, и (3) освещение, пригодное для фотосинтеза.
Растения, выращенные в темноте, «стараются» повысить чувствительность к свету, повышая содержание фитохрома А (именно Ф(А) отвечает за восприятие света любого качества). При освещении (независимо от того, К или ДК), концентрация Ф(А) снижается примерно в 100 раз.
Киназная активность фитохромов зависит от состояния хромофора. Форма Ф730 является активной (она получила красный свет с λ = 660), а Ф660 (образуется в темноте или на ДК) не активна как киназа. Фосфорилирование фитохромов также меняет их активность. Возможно, фитохромы могут автофосфорилироваться. В число возможных вторичных мессенджеров фитохромного сигнала входят гетеротримерные G-белки, циклическая ГМФ и кальций.
У растений, выращенных в темноте, фитохромы обычно находятся в цитозоле. Для перемещения фитохромов А и В в ядро нужны разные световые стимулы. Так, в конформации Ф660 фитохром В не может пересекать ядерную мембрану, а после фотоконверсии красным светом аккумулируется в ядре. Ф(А) переходит в ядро при освещении как ДК высокой интенсивности, так и красным светом. Интересно, что на С-концевом участке у фитохромов есть два PAS-домена*, которые встречаются в транскрипционных факторах. PAS-домены важны для ДНК-белковых и для белок-белковых взаимодействий. Повреждение PAS-доменов нарушает функционирование фитохрома. Это означает, что, проникая в ядро, фитохром участвует в геномной регуляции, связываясь в комплексы с факторами транскрипции.