Жасуша патофизиологиясы
Жасуша бүліністерінің этиологиясы.Жасуша бүліністеріне көптеген себепкер ықпалдар әкеледі. Оларды сырттан әсер ететін- экзогендік және организмнің өзінде пайда болатын – эндогендік– деп екі үлкен топқа ажыратады. Экзогендік бүліндіргіш ықпалдар өз алдына инфекциялық және бейинфекциялық болып бөлінеді. Оларға:
●биологиялық ықпалдар (микробтар, вирустар, қарапайым жәндіктер, майда саңырауқұлақтар, аутоантиденелер мен цитотоксиндік иммундық глобулиндер т.б.);
●физикалық ықпалдар (иондағыш сәулелер, ыстық немесе суық температуралар, электр соққы, электромагниттік толқындар т. б.);
●механикалық ықпалдар (жарақат, жаншылу, қысылу т.б.);
● химиялық ықпалдар (органикалық немесе бейорганикалық улы заттар, кейбір дәрі-дәрмектер, гипоксия, реоксигенация т.б.)
● психогендік ықпалдар ( жан жарақаты, рухани күйзелістер, ауыр эмоциялар т.б.) – жатады.
Эндогендік себепкер ықпалдар туа біткен және жүре пайда болған болып бөлінеді. Туа біткен себепкер ықпалдарға жасуша мембранасындағы тұқым қуалайтын ақаулардың болуы жатады. Бұл кезде мембрананың құрамында кейбір нәруыздар, фосфолипидтер немесе ферменттер болмауы ықтимал.
Жүре пайда болған эндогендік себепкер ықпалдарға:
● жүйкелік эндокриндік реттелулердің біріншілік бұзылыстары;
● иммундық бұзылыстар немесе аллергиялық үрдістер;
● зат алмасу өнімдерінің (Н+, К+, Са2+, асқын тотықтар т. б.) жиналып қалуы;
● организмде су мен электролиттер алмасуының бұзылыстары;
● бүйрек қызметінің жеткіліксіздігі;
● ағза қызметінің тым қатты көтерілуі немесе ұзақ мерзім болмауы;
● жасушаның қартаюы т. т. - жатады.
Жасушаның бүліністері пайда болуында әсер ететін ықпалдармен бірге, жасушаның өзінің ішкі жағдайы да маңызды. Жасушаның алғашқы жағдайына байланысты кез келген ықпал қорғаныстық немесе бүліндіргіш әсер етуі мүмкін.
Жасушаның жағдайлары тұқым қуалайтын қасиеттерімен анықталады және жүре пайда болған авитаминоздар, созылмалы гипоксия, зат алмасу үрдістерінің өзгерістері т. т. жасушаның көптеген әдеттегі әсерлерге төзімділігін төмен-детеді. Гормондық реттелудің бұзылыстары зат алмасу үрдістерін бірінші бұзып, сол арқылы әртүрлі әсерлерге жасушаның тұрақтылығын азайтады. Мәселен, гипертиреоз кезінде оның гипоксияға төзімділігі төмендейді. Қантты диабет кезінде микробтарға төзімділігі азаяды.
Сонымен бірге, көптеген (биологиялық, химиялық, физикалық) ықпалдар жиі бірігіп әсер етуі мүмкін.
Жасуша бүліністерінің жалпы даму жолдары мына бұзылыстардан байланысты болады:
●энергиямен қамтамасыз етілуінің бұзылыстарына;
● жасуша мембраналарының бұзылыстарына;
●гендік құрылымдарыныңбұзылыстарына;
●жасушада иондар мен сұйықтардың алмасуы бұзылыстарына;
●жасуша қызметтерінің реттелулерінің бұзылыстарына - байланысты дамиды.
Жасушаның энергиямен қамтамасыз етілуінің бұзылыстары мына жағдайларда байқалады:
● макроергиялық фосфорлық қосындылардың (АТФ) түзілуі бұзылудан; Ол өз алдына мына жағдайларда:
♣ оттегінің жетіспеушілігінен;
♣ қоректік заттардың аздығынан;
♣ гликолиздік және тотығу-тотықсыздану ферменттерінің белсенділігі төмендеуінен;
♣ митохондрийлардың бүліністерінен – байқалады.
● энергияның жасуша ішінде тасымалдануы бұзылыстарынан; Мәселен, адениннуклеотидтрансфераза ферменті митохондрийлардың ішінен олардың ішкі мембранасы арқылы АТФ энергиясын шығарады. Ары қарай оны митохондрийдің мембранасындағы креатинфосфокиназа (КФК) ферменті креатинге ауыстырып, креатин фосфат (КрФ) құрады. Креатин фосфат жасушаның цитоплазмасына түседі де, қызмет атқаратын құрылымдардың КФК ферментінің көмегімен оның бір фосфор қышқылы АДФ молекуласына ауыстырылып, АТФ түзіледі. Ол жасушаның тіршілікке қажетті міндетін атқаруға пайдаланылады. Осындай энергияны тасымалдау жүйесі көптеген бүліндіргіш ықпалдардан бұзылады.
● жасушаның энергияны пайдалануы бұзылыстарынан – дамиды.Әрбір жасуша өзі өндірген АТФ қуатын ғана пайдаланады, басқа жасушаларда өндірілген немесе емдік мақсатта сырттан енгізілген АТФ пайдаланылмайды. Энергияны пайдалану әртүрлі уыттардың әсерлерінен АТФ-аза ферменттерінің белсенділігі төмендегенде байқалады.
Жасуша мембраналарының бұзылыстары.
Биологиялық мембрана екі қабат фосфолипидтердің молекуласынан және нәруыздардан құрылады (8-сурет). Атқаратын міндеттемелері бойынша нәруыздар әртүрлі болады. Оларға иммундық және құрылымдық нәруыздар, иондар үшін каналдар құратын нәруыздар, медиаторлар мен гормондардың рецепторлары, ферменттер, оның ішінде тасымалдағыш ферменттер жатады.
Мембрана мен өзара арақатынастарына қарай нәруыздар липидтік екі қабатпен өте тығыз байланысқан және мембрананы бір немесе бірнеше рет жарып өтетін тұтастық (интегралдық) және липидтік екі қабаттың бір бетімен ғана байланысқан шеткері нәруыздар болады. Тұтастық нәруыздарды бөліп алғанда мембрананың тұтастыгы бұзылады. Шеткері нәруыздар тіпті әлсіз әсерлерден (осмостық қысым, ортаның қышқылдығы, Са2+ иондарының мөлшері өзгергенде) мембранамен байланысын жоғалтады. Мембрананың құрылымдық тұтастыгы липидтермен анықталады. Жануарлар жасушаларының қабықтарында фосфолипидтер басым болады жөне липидтердің құрамына холестерин кіреді.
return false">ссылка скрытаМембранада көмірсулары бос күйінде кездеспейді. Олар көбінесе нәруыздармен, липидтермен байланысқан түрде болады және әртүрлі қызметтер атқарады. Мысалы, көмірсулар тізбегі нәруыздармен байланысып, мембрананың қажетті бөлшегіне олардың байланысуын қамтамасыз етеді. Май қышқылдары мембранада глико- немесе фосфолипидтердің құрамында болады. Мембранадағы липидтік екі қабатта бірнеше түрлі қозғалыстар болады: тербелулік, айналмалы, мембрана бойынша көрші молекулалармен орын ауыстыру арқылы, мембрананың бір жағынан екінші жағына өту (флип-флоп қозғалысы). Бірінші үш қозғалыс минөтіне миллион рет болса, соңғысы сағатына 1 рет немесе одан да сирек болады. Сонымен мембрана көптеген әлсіз байланыстармен бекітілген көп қозғалатын жүйе. Кейбір байланыстар жеңіл үзіліп, жаңадан пайда болып жатады. Көпшілік мембраналардың арасында жасушаның сыртқы беті маңызды орын алады. Ол гликопротеидтерден тұрады. Гликопротеидтер жасушаның иммундық қасиетін анықтайды. Жасушаның сыртқы бетінің өзімен түйісетін басқа жасушалар мен молекулаларды (гормондарды, медиаторлар, цитокиндерді т. б.) танитын ғажап қасиеті бар. Ол цитоплазмалық мембрананың сыртындағы гликопротеидтермен қамтамасыз етіледі. Тану қызметі тіннің жасуша аралық өзара әсерлерінің, лимфоциттердің иммундық жауаптарының, гормондар мен жасуша ішіндегі үрдістердің реттелуінің, медиаторлардың әсерлерінен жасушалардың қозуының негізінде жатады. Арнайы қызмет атқаратын жасушаларда биологиялық мембраналары көптеген құрылымдар құрады. Мысалы, бауыр жасушаларының мембраналары организмде улы қосындыларды усыздандыруға жауапты эндоплазмалық торшалардың күрделі торын құрады. Без жасушаларының мембраналары гормондардың түзілуіне жауапты көпіршіктер мен өзектердің торын қалыптастырады және Гөлдж жүйесін құрастырады. Жүйке жасушаларында биологиялық мембраналар бір жасушадан екіншісіне сыртқы хабарды тасымалдау, тарату және қабылдау қызметтерін атқарады. Бұлшықет жасушаларының мембранасы (сарколемма), Т-жүйесі деп аталатын, жіңішке түтікшелер ретінде, жасушаның ішіне кіреді. Ол жасуша ішіндегі бұлшықет қысқаруын қадағалайтын саркоплазмалық торлармен қалыптасқан көпіршіктермен жалғасады. Эпителий жасушаларында биологиялық мембраналардың қызметі жасушаның қай жағында орналасқанына байланысты болады. Тіндер жағына қараған мембраналар иондар мен заттарды белсенді тасымалдау қызметін атқарады. Сыртқа қарайтын үстіңгі жағындағы мембраналар енжар тосқауыл қызметін орындайды.
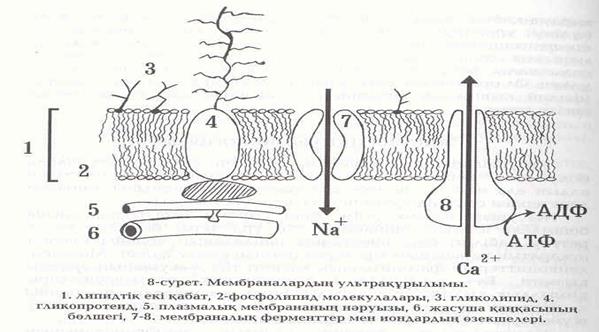
Биологиялық мембраналар жасуша іші мен сырты арасында заттар мен иондардың белгілі дәрежеде бөлінуін ұстап түрады. Жасуша ішінде К+ иондары оның сыртындағы деңгейінен бірнеше есе артық. Керісінше, натрий иондары жасуша сыртында оның ішіндегі деңгейіне қарағанда он есе көп. Сонымен қатар саркоплазмалық торшалардың көпіршіктерінде кальций иондарының мөлшері жоғары болады. Иондардың жасуша іші мен сыртында біркелкі бөлінбеуі мембранадағы липидтік екі қабатта иондық «насостардың» немесе «помпалардың» болуымен қадағаланады. Олардың қызметтері АТФ-ның ыдырауынан пайда болатын энергияны пайдалану арқылы орындалады. Иондық «насостардың» негізгі үш түрі белгілі: Н+ - АТФаза, Са2+- АТФаза, Nа+, К+ - АТФаза. Біріншісі 1 молекула АТФ ыдырағанда 2 атом Н+ тасиды, Са2+ АТФаза - 1 атом Са2+, Na+ -, К+ - АТФаза 3 атом Nа+ және 2 атом К+ тасиды. Бұл насостар жасушаның әртүрлі мембраналарында орналасқан. Бірақ Н+-АТФаза митохондрийлардың, Са2+- АТФаза саркоплазмалық ретикулумның, Nа+, К+ - АТФаза цитоплазмалық мембраналарда көбірек болады. Бұл АТФазалар Н+, Са2+, Nа+, К+ - иондарының бір келкі бөлінбеуін қадағалайды. Саркоплазмалық ретикулумнан және жасуша сыртынан оның ішіне Са2+ иондары тез арада кіруі еттің қысқаруына және жасушада нәруыз түзілуінің көтерілуіне әкеледі. Осы кезде саркоплазмалық ретикулум мембранасында Са2+-АТФ-аза әсерленіп, Са2+ иондарын саркоплазмадан сорып алады. Осының нөтижесінде ет талшықтарының босаңсуы болады және нәруыздардың түзілуі азаяды. Н+-АТФ-аза митохондрийларда тотығулық фосфорлануды арттыру арқылы АТФ түзу үшін қажет. Бұл үрдіс митохондрийлардың ішкі қабығы астында өтеді. Онда тотығатын заттардан молекулалық оттегіге дейін электрондарды таситын ферменттердің бір тұтас жүйесі бар. Осылардың нәтижесінде түзілген энергия тіршілікке қажетті қызметтер орындалуы үшін пайдаланылады. Егер Са2+ иондары не бір себептерден жасуша ішіне артық түссе, онда митохондрийлар кальцийді сорып алады. Бұның бір жағынан икемделістік маңызы бар. Өйткені ол бұлшықет талшықтарын сіресіп жиырылып қалудан сақтандыруға бағытталған. Екінші жағынан митохондрийларда Са2+ иондарының жиналып қалуы, тотығу-тотықсыздану үрдістерінің бұзылуына, энергияның аз түзілуіне әкеледі.
МЕМБРАНА БҰЗЫЛЫСТАРЫНЫҢ ДАМУ ЖОЛДАРЫ (ПАТОГЕНЕЗІ).
Биологиялық мембраналардың бүліністері мына 4-жолмен дамуы ықтимал:
● мембранадағы еркін радикалды тотығу мен майлардың асқын тотығуынан;
● фосфолипаза, липаза, протеаза ферменттерінің артық әсерленіп кетулерінен;
● мембраналардың механикалық (осмостық) бүліністерінен;
● иммундық (аллергиялық) әсерлерден.
ЕРКІН РАДИКАЛДЫ ТОТЫҒУ МЕН МАЙЛАРДЫҢ АСҚЫН ТОТЫҒУЫ.
Майлардың асқын тотығуы (МАТ) – деп жасуша мембраналырының фосфолипидтерінің құрамына кіретін қанықпаған май қышқылдарының еркін радикалды тотығуын айтады. Оның тым артып кетуіне бос радикалдар (супероксиданионрадикал - О‾, гидроксил радикалы – ОН •, сутегінің радикалы - Н • , сутегінің асқын тотығы – Н2О2 , синглетті оттегі - 1О) әкеледі. Организмде еркін радикалды тотығу белгілі деңгейде қалыпты жағдайларда болып тұратын үрдіс. Ол электрондардың тасымалдануына, тыныстық ферменттердің тізбектерінде, простагландиндер мен лейкотриендер түзілуіне, жасушалардың өсіп-өнуі мен нақтылануларында, фагоцитозда, катехоламиндердің алмасуларында т.б. құбылыстарда өте маңызды орын алады. Осы жағдайларда оттегінің молекуласы бір электрон қосып альш, супероксиданионрадикалга (О ‾ ) ауысады. Қалыпты жағдайда супероксиданион радикалы фермент супероксиддисмутазаның (СОД) әсерімен сутегі асқын тотығына (Н2О2), ары қарай Н2О2 каталаза мен глутатионпероксидазаның әсерінен суға және оттегіге айналады.
Көрсетілген ықпалдардың әсерлерінен О ‾ - ның тым артық өндірілуінен антиоксиданттық ферменттермен жойылып үлгермейді де, супероксиданион радикалы мен сутегінің асқын тотығынан өте озбыр гидроксил радикалы (ОН • ) пайда болады. Ол өз алдына қанықпаған май қышқылдарымен байланысып, олардың гидроксилді радикалдарын және асқын тотықтарын (RОН°; RООН°) тудырады. Олар ары қарай диенді қоспалардың, малондық диальдегидтердің және Шифф негіздерінің пайда болуына әкеледі. Қалыпты жағдайларда бұл асқын тотығудың өнімдері табиғи антиоксиданттармен (альфа-токоферол, глутатион, аскорбин қышқылы, коэнзим-Q т. с. с.) байланыстырылады да өздерінің белсенділігін жояды. Ал, антиоксиданттар жүйесінің тұқым қуатын немесе жүре пайда болған жеткіліксіздіктері кездерінде бос радикалдардың бейтарапталуы болмайды да, майлардың асқын тотықтары май қышқылдарының жаңа молекулаларымен әрі қарай тізбек тотығу реакциясына ілігеді.
Сонымен, жасуша мембраналарының бүліністерінде майлардың асқын тотығуы екі түрлі жағдайларда болуы ықтимал:
● біріншісі, бос радикалдардың тым артық өндірілуінен ;
● екіншісі, антиоксиданттық жүйелердің жеткіліксіздігінен.
Осыған байланысты организмде белсенді бос радикалдар артық өндірілуі мына жағдайларда:
♣ организм улы заттармен (ауыр металдармен, төрт хлорлы көміртегімен т. б.) уланғанда;
♣ механикалық жарақат, жан-дүниелік күйзелістер т. б. ауыртпалықтар кездерінде катехоламиндерден бос радикалдар құрылғанда;
♣ иондағыш сәулелермен сәулеленгенде;
♣ оттегімен уланғанда;
♣ тіндер қабынуға ұшырағанда;
♣ фагоцитоз кезінде;
♣ гипоксия және реоксигенация кездерінде – тым артып кетеді.
Антиоксиданттық жүйелердің туа біткен және жүре пайда болған жеткіліксіздігі мына жағдайларда:
● антиоксиданттық ферменттердің (супероксиддисмутаза, каталаза, глютатионпероксидаза т.б.) түзілуі бұзылыстарында;
● бұл ферменттердің қызмет атқаруларына қажетті темір, мыс, селен иондарының тапшылығында;
● антиоксиданттық витаминдердің (витамин Е, С т.б.) жетіспеушілігінде;
● зат алмасуларының пентоздық-фосфаттық жолдарының бұзылыстары- нан антиоксиданттарды тотықсыздандыруға қажетті НАДФН және НАДН молекулалары жеткіліксіз түзілгенінде - байқалады Ауаны ластайтын химиялық уытты газдар мен заттар (ауыр металдар, фосфор, гербицидтер мен пестицидтер т.б.) антиоксиданттық ферменттердің белсенділігін төмендетуі және антиоксиданттық витаминдерге организмнің мұқтаждығын арттыруы мүмкін. Бұл кездерде болмысты зат алмасулары кездеріндегі өндірілген бос радикалдар бейтарапталмай майлардың асқын тотығуын өршітіп жібереді.
Қай жолмен дамығанына қарамай, майлардың асқын тотығуы жасуша мембраналарының барлық қызметтерінің бұзылыстарына әкеледі.
ФОСФОЛИПАЗА, ЛИПАЗА, ПРОТЕАЗА ФЕРМЕНТТЕРІНІҢ АРТЫҚ ӘСЕРЛЕНІП КЕТУІ.
Бұлардың әсерленуі жоғарыда көрсетілген себепкер ықпалдардан болады. Сонымен бірге:
● биологиялық ықпалдар (жұқпалар, жылан және ара улары т. б.);
● ишемия;
● Са2+ иондарының жасуша ішінде артық жиналуы фосфолипаза А мен липазаны, қышқыл протеазаларды қатты әсерлендіреді.
Осыдан мембраналарда фосфолипидтер, майлар, гликолипопротеидтер ыдырайды, босаған қанықпаған май қышқылдары асқын тотығуға ілігеді және артық бос май қышқылдарының өздері мембранаға тікелей бүліндіргіш әсер етеді. Сонымен бірге, фосфолипаза А2 –ферментінің әсерінен фосфолипидтерден босаған арахидон қышқылынан простагландиндер мен лейкотриендер түзіледі. Фосфолипидтердің құрамынан май қышқылдары бөлініп алынудан олардың құйыршықтарында май қышқылынан тұратын бір ғана тізбек сақталады. Осындай фосфолипидтерді лизофосфолипидтер дейді. Олар мембранаға одан сайын бүліндіргіш әсер етеді. Ишемия кезіндегі жасушалардың бүліну жолдары 18-сызбанұсқада келтірілген.
18- сызбанұсқа
Ишемия кезіндегі жасушалардың бүліну жолдары

МЕМБРАНАЛАРДЫҢ МЕХАНИКАЛЫҚ (ОСМОСТЫҚ) БҮЛІНІСТЕРІ.
Механикалық жарақаттар, сыртынан ауыр заттармен қысылып қалуы мембрананың жыртылып кетуіне т.с.с. бүліністеріне әкеледі. Организмде су мен электролиттер алмасуы бұзылудан мембраналардың осмостық керіліп созылуы, содан механикалық жыртылуға дейінгі өзгерістер байқалады. Бұл кезде жасуша ішіндегі осмостық қысымның жасуша сыртындағы деңгейінен артық болуынан су көптеп оның ішіне кіреді. Мәселен, бүйрек қызметінің жеткіліксіздігі кезінде денеде жиналған артық су жасушалардың ішіне еніп, олардың ісінуіне әкеледі Осыдан жасуша мембраналарының керіліп созылуынан эритроциттердің еріп кетуі байқалады. Жасушалардың осмостық бүліністері гипотониялық гипогидратация мен гипергидратация кездерінде болады Керісінше, жасуша ішінде осмостық қысым сыртындағыдан төмен болғанда жасушаның бүрісіп қалуы байқалады. Содан жасушаның құрылымы мен атқаратын қызметтері бұзылады (Су мен электролиттер алмасуының бұзылыстарын қараңыз).
ИММУНДЫҚ (АЛЛЕРГИЯЛЫҚ) ӘСЕРЛЕР.
Антиденелер мен сезімталдығы көтерілген лимфоциттер, комплементтің құрамбөлшектері, цитокиндер, табиғи жендет жасушалар, макрофагтар жасуша мембраналарына бүліндіргіш әсер етеді. Макрофагтар бүлінген жасушалардың бөлшектерін жеп қояды. Фагоцитоздық қызметтің негізінде лейкоциттерде еркін радикалды тотығудың артуы, оттегінің бос радикалдары өндірілуінің көбеюі маңызды орын алады.
Сонымен бірге, нысана-жасушалардың бүліністері антиген-антидене кешендерінің, комплемент құрамбөлшектерінің ( С3а, С5в, С8, С9 т.б.), цитотоксиндік Т-киллер және табиғи жендет (киллер) жасушалардың, өспені жоятын фактордың, гамма-интерферонның т.с.с. цитокиндердің, лизосомалық ферменттердің, оттегінің бос радикалдарының әсерлерінен дамиды (Аллергия тақырыбын қараңыз).
Биологиялық мембраналардың қандайда болмасын жолмен бүлінуі олардың барлық қызметтерінің бұзылыстарына әкеледі.
Мембрананың тосқауылдық қызметінің бұзылыстары.
Қанықпаған май қышқылдары асқын тотығып кетуден мембрананың тұтқырлығы көтеріледі, қимылдық қозғалыстары азаяды. Асқын тотығулық «кластерлер» деп аталатын, мембранада тесіктер пайда болады. Осыдан мембрананың өткізгіштігі көтеріледі, жасуша ішінде және сыртындағы иондардың қалыпты арақатынасы өзгереді, иондық дисбаланс дамиды. Жасуша ішінен калий иондары сыртқа шығып, натрий иондары ішіне түседі. Натрий иондарымен бірге су енеді, сөйтіп жасуша ішінде ісіну пайда болады. Бұлшықет жасушаларының ішіне көптеп Са2+ иондары кіруден олардың сіресіп қалуына әкелетін қатты жиырылулары болады. Жасуша іші мен сыртында иондардың арақатынасы бұзылудан қозымды тіндерде мембраналық потенциал, әрекеттік потенциал бұзылады. Осыдан миокард жасушалары бүлінгенде ЭКГ-де, ми нейрондары бүлінгенде ЭЭГ-да, бұлшықет жасушалары бүлінгенде электромиографияда (ЭМГ) өзгерістер пайда болады.
Сонымен бірге Са2+ иондары кәлмодулинмен байланысып, фосфолипаза, қышқыл протеаза ферменттерін әсерлейді. Осылардың нәтижесінде кері айналып соғу шеңбері бойынша жасуша мембранасының қызметтері бұзылуын одан сайын күшейтеді. Мембрана өткізгіштігі көтерілуіне байланысты жасуша ішіндегі ферменттер сыртына шығып кетеді, жасуша аралық сұйықтармен қанға түседі. Мәселен, бауыр, жүрек жасушалары мембраналарының өткізгіштігі жоғарылауынан қанда аспартатаминотрансфераза, алатаминотрансфераза, лактатдегидрогеназа, креатинфосфокиназа т. б. ферменттердің деңгейлері көтеріледі. Бұлшықет жасушалары мембраналарының өткізгіштігі көтерілуінен қанда миоглобин көбейеді. Бұлардың қандағы деңгейін анықтау клиникада дертті аңғару үшін кеңінен қолданылады.
Мембраналардың рецепторлық қызметтерінің бұзылуы.Мембраналарда көптеген рецепторлар (қабылдағыштар) болады. Бұларға әсер ететін заттарды лигандалар деп атайды. Оларға нейромедиаторлар, цитокиндер, пептидтік гормондар, антигендер, стероидтық және тиреоидтық гормондар т.с.с. заттар жатады.Лигандалардың әсерлерінен рецепторлар қоздырылып, екінші қатардағы дәнекерлер өндіріледі де жасуша қоздырғышқа жауап қайтарады.
Мембраналық рецепторлар жасушаға әсер еткен дабылды қабылдап, оны жасуша ішіне қабарлайды. Осыдан жасуша ішіндегі химиялық үрдістерді реттеуге қатысатын екіншілік дәнекерлер өндіріліп, жасушаның қызметтері атқарылады. Бұл екіншілік дәнекерлерге цАМФ, цГМФ, инозитолтрифосфат, диацилглицерол, Са2+ -иондары жатады.
Рецептордан жасушаның ішіне берілетін хабардың негізгі жолының бірі болып, мембраналық аденилатциклаза ферментінің әсерленуі есептеледі. Осының нөтижесінде АТФ молекуласынан циклдік АМФ түзіледі. Ол өз алдына протеинкиназа ферменттерін әсерлеп, фосфорлау үрдістерін күшейту арқылы көптеген жасуша ішіндегі үрдістерді сергітеді (9-сурет). Осыдан мембранада белсенді тасымалдау, нәруыздардың түзілуі, ферменттердің әсерленуі күшейеді де, жасушаның биологиялық қызметі көтеріледі. Мысал ретінде фосфорила-заның әсерлену жолын келтіруге болады. Глюкагон немесе адреналиннің әсерлерінен аденилатциклазаның белсенділігі көтеріліп циклдік АМФ түзіледі. Ол өз алдына протеинкиназа ферментін әсерлеп, фосфорилаза В-ны фосфорилаза А-ға айналдырады. Соңғысы жасушадағы гликогенді ыдыратып, глюкозаның мөлшерін көбейтеді. Әрі қарай глюкозаның тотығуы нәтижесінде тіршілікке қажетті макроэргиялық қосындылар түзіледі. 1 моль глюкоза тотығуынан 40 моль АТФ өндіріледі.
Рецепторлардан жасушаның ішіне берілетін хабардың тағы бір маңызды жолы болып Са2+ иондары арқылы өтетін үрдістер есептеледі (9-сурет).
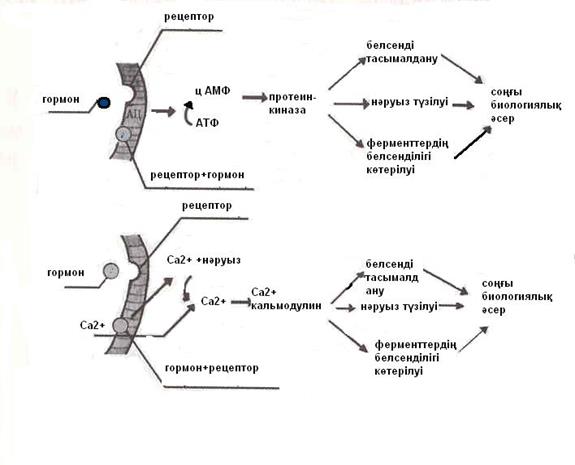
9-сурет. Жасуша ішіндегі өтетін үрдістерді реттеу жолдары.
Мәселен, гормондар жасуша мембранасындағы рецепторлармен байланысып, Са2+ иондарының жасуша ішіне енуін күшейтеді. Осының нәтижесінде жасуша ішінде Са2+ иондары көбейеді. Са2+ иондары кәлмодулинмен байланысып фосфорилазаны әсерлейді. Артынан гликогеннің ыдырауы артып, глюкозаның деңгейі көтеріледі. Сонымен бірге, жасуша ішіне хабар жеткізілуіне фосфатидилинозитолдың өнімдері қатысады. Олар G-нәруыздарымен бірігіп әсер етеді. Мембранадағы инозитолфосфатаза (фосфолипаза С) әсерленуінен фосфатидилинозитол-4,5-дифосфат инозитолүшфосфатқа және диацилглицеринге ыдырайды. Бұлар жасуша ішінде Са2+ - иондарын көбейтіп, протеинкиназа С –дің белсенділігін көтереді. Осыдан көптеген жасуша ішілік нәруыздардың фосфорлануы болады, жасуша құрылымына қажетті нәруыздардың әсерленуі, олардың өсіп-өнуін қадағалайтын гендік бағдарламаның әсерленуі байқалады, простагландердің түзілуі артады.
Бұл көрсетілгендердің барлығы цитоплазмалық мембрананың жағдайына байланысты. Мембрананың қанықпаған май қышқылдарының асқын тотығуға ілігіп кетуінен микротұтқырлығы көтерілуі рецепторлардың липидтік екі қабаттың бойымен қозғалуын шектейді. Мембрананың рецепторлық қызметтері бұзылуы нәтижесінде жасушалардың нейрогуморалдық реттеу хабарларына жауабы өзгереді, хабардың жасуша ішіне берілуі бұзылады. Осыдан жасушада заттардың алмасулары және оның қызметі өзгереді.
Жасушалардың бірін-бірі тану үрдістерінде және олардың өзара әсерлерінде мембрана бетінде орналасқан хабаршы молекулалардың маңызы үлкен. Оларға тіндік үйлесімділік, топтық ажыратулық, тінге арнайылық т. б. антигендер жатады. Бұлардың құрылымы мен ерекшеліктерінің өзгеруі жасуша-аралық әсерлердің бұзылуына және қажетсіз жасушалардың жойылуына әкеледі. Мысалы, бүлінген жасушалар сыртындағы гликопротеидтерді жоғалтады. Көптеген ауру туындататын микроорганизмдер (тырысқақ вибрионы, тұмаудың вирусы т. б.) нейраминидаза ферментін шығарып, жасуша қабығындағы антигендік қасиеті бар, жасырын бөлшектерін ашады. Бұл бөлшектер қалыпты аутоантиденелермен байланыса алады және организмнің иммундық жауабын шақырып, артынан бүлінген жасушаларды жоятын иммундық әсерлердің (комплемент, макрофагтар) шабуылына ілінеді. Осылай олар организмнен аластанады. Мембрана өзіне вирустық нәруыздың бөлшектерін бекітеді. Осыдан бүлінген жасушаларды жендет жасушалар тез байқап, жойып отырады.
Жасуша рецепторларының бұзылыстары мына жағдайларда болуы ықтимал:
● рецепторлардың сезімталдығы өзгеруден;
● рецепторлардың саны өзгеріп кетуден;
● рецепторлық нәруыздардың конформациялық бұзылыстарынан;
● рецепторлардың айналасын қоршаған липидтердің өзгерістерінен. Бұл өзгерістердің барлығы мембрананың бұзылыстарында байқалады.
Екіншілік дәнекерлердің ауытқулары да жасуша ішілік үрдістердің реттелулерінің бұзылыстарына әкеледі. Мәселен, жүрек ет жасушаларында цАМФ жиналып қалудан, кардиомиоциттерде мембраналық потенциал қалыптасуы бұзылып, жүрек аритмиялары дамиды. Бронхиалық демікпе кезінде кеңірдекшелердің тегіс еттерінде цАМФ азайып кетуден, олар қатты жиырылып қалады. Саркоплазмалық торшалардың Са 2+ -АТФ-аза ферментінің белсенділігі төмендеуден Са2+ -иондарының бұлшықет жасушаларының іштерінде жиналып қалуы, олардың сіресіп жиырылуына әкеледі.
Мембраналардың насостық қызметтерінің бұзылуы.Жасуша сыртындағы мембрананың Nа+-, К+-АТФаза ферментінің белсенділігі төмендейді. Сондықтан жасуша ішінде жиналған натрий иондары сыртқа шығарылып, калий иондарының оның ішіне кіруі бұзылады. Саркоплазмалық ретикулум мембранасында Са2+-АТФаза ферментінің белсенділігі төмендейді. Сөйтіп Са2+ иондары саркоплазмада жиналып қалады. Мембранадағы насостардың әлсіреуі май қышқылдарының асқын тотықтарының (RОН°) әсерлерінен болатыны белгілі. Сонымен бірге олардың бұзылыстарында энергия жеткіліксіздігінің де маңызы үлкен (төменде келтірілген).
Мембраналардың каналдық қызметтерінің бұзылуы.Мембранадағы липидтердің асқын тотығуынан және фосфолипаза, липаза ферменттерінің артық әсерленулерінен, фосфолипидтер ыдыратылады. . Са2+- тәуелді протезалардың ықпалдарынан канал құратын нәруыздар құрылымдық өзгерістерге ұшырайды. Осыдан және бұл нәруыздардың молекулаларында амин қышқылдарының кеңістікте орналасу тәртіптерінің өзгерістері нәтижесінде мембранадағы каналдардың қызметтері бұзылады
Мембраналардың қызметтерінің бұзылыстары, жасушаның сыртқы мембранасында ғана болмай, жасуша ішіндегі құрылымдардың мембраналарында да болады. Осыған байланысты митохондрийлардың мембраналары бұзылудан энергия түзілуі бүлінеді. Қалыпты жағдайда тотығу-тотықсыздану реакциялары нәтижесінде босаған энергияның бір бөлшегі жылу ретінде тарайды, ал қалғаны (60-%-ға жуығы) энергиялық қосындылар (АТФ, КрФ) түрінде жиналады. Соңғылары артынан тіршілікке қажетті организмнің әртүрлі қызметтерінде (ет жиырылу, бездердің сөлденісі, ойлау т. с. с.) пайдаланылады. Митохондрийлардың мембраналары бұзылыстарынан онда су, Са2+ иондары, липофуксин, ферритин т.с.с. заттар артық жиналып қалады. Сонымен бірге олардағы ферменттер жүйесі әлсірейді. Сондықтан митохондрийларда тотығу мен фосфорланудың ажырауы болады да макроэргиялық қосындылар аз түзіледі. Осыдан организмде энергияға деген тапшылық жағдай дамиды, мембраналардағы насостардың қызметтері одан сайын әлсірейді. Мембраналардың бүлінуі нәтижесінде митохондрийлар ыдырайды, ериді немесе ісінеді, олардың тарақшалары (кристалары) бөлшектенеді, жоғалады. Митохондрийлардың ішінде бос қуыстар (вакуолдар) пайда болады.
Жасуша ішіндегі ядроның мембраналары бұзылыстарынан гендік ақпараттың сақталуы мен туынды жасушаларға берілуі, нәруыз түзілуі бұзылады. Ядрода ДНҚ молекуласының репликациясы, РНҚ-ның транскрипциясы өзгереді. Жасушалардың әртүрлі бүліністерінде ядрода хроматиннің тығыздығы, құрылымы және ядро ішінде тарауы, ядро түрінің өзгеруі болады. Бұл кезде ядроның ісінуі, оның жиырылып, бүрісіп қалуы (пикноз), жыртылуы және ыдырауы (кариорексис және кариолиз) байқалады. Ядродағы ядыршықтардың саны мен құрылымы өзгереді.
Ядроның бүлінуімен бірге жасуша ішіндегі кедір-бүдыр эндоплазмалық торшалардың, рибосомалар мен Гөлдж кешендерінің мембраналары, бұзылыстарына байланысты жасушаларда нәруыздар, ферменттер, гормондар түзілуі де бүлінеді. Тегіс эндоплазмалық торшалардың мембраналары бұзылуынан жасуша ішінде үшглицеридтердің, май қышқылдарының, стероидтардың, фосфолипидтердің түзілуі бүлінеді.
Лизосомалардың мембранасы бүлінуі нәтижесінде гидролиздік ферменттер (қышқыл фосфатаза, ДНК-аза, РНК-аза, катепсин Д т. б.) босап шығады. Олар айналасындағы жасушаларды ыдыратып, тіршілігін жояды.
Гендік құрылымдарының бұзылыстары.
ДНК молекуласының құрылымы бұзылуы немесе оның ыдырап кетуі жасуша тіршілігінің жоғалуына жиі әкеледі. Оның бүліністері мына себептерден болуы ықтимал:
● ДНК молекуласына өте күшті иондағыш сәулелер, бос радикалдар, майлардың гидроасқын тотықтары т.б. әсер етулерінен оның құрамбөлшектері бүлінеді;
● нуклеаза ферменттері қатты әсерленіп кеткенде ДНК ыдырайды;
● белсенділігі қатты көтерілген трансфераза ферменттерінің әсерлерінен нуклеотидтердің арасындағы байланыстар үзіліп, ДНК молекуласын ыдыратады.
Осыдан:
♣ гендердің мутациясы;
♣ дерттік гендердің (мәселен, онкогендердің, апоптоздық гендердің) әсіреленіп кетуі;
♣ тіршілікке маңызды (мәселен, ферменттердің түзілуін қадағалайтын) гендердің белсенділігі төмендеп кетуі;
♣ геномға бөтен текті ДНК (мәселен, вирустың ДНК-сы) еніп кетуі (трансфекция);
♣ бүлінген ДНК молекуласының қалпына келуі бұзылуы – тектік ақпараттың бұзылыстарына әкеледі. Бұндай өзгерістер сау жасушаның өспе жасушасына айналуында маңызды орын алады.
Жасуша қызметтері реттелулерінің бұзылыстары.
Тін жасушалары жүйкелік-эндокриндік, паракриндік және аутокриндік жолдармен реттелінеді. Осыған байланысты ағза мен тіннің жүйкеленуі бұзылғанда жасушаларда дистрофия дамып, мүшенің семіп қалуы болады. Ішке сөлденістік бездердің гормондарының тым артып кетуінен немесе олардың болмауынан жасушалардың дамуы, жетілуі және нақтылануы бұзылады. Паракриндік реттелу – деп көршілес орналасқан жасушалардың әртүрлі гормондық заттар, цитокиндер, өсу факторларын т.б. кесек молекулалар бөліп шығарып, нысана жасушалардың өсіп-өнуін, жетілуі мен нақтылануын, өзіне тән міндеттемелерін атқаруын реттеуді айтады. Жасуша мембраналарындағы оларды қабылдайтын рецепторлардың бұзылыстарынан паракриндік реттелу өзгереді.
Аутокриндік реттелу қатерлі өспе жасушаларына тән құбылыс. Өспе жасушалары жоғарыда келтірілген реттелу жолдарына бағынбай өзбеттерінше өзін-өзі реттеп өседі (Өспе өсуін қараңыз).
Жасуша бүліністерінің көріністері.
Жасуша бүліністерінің көріністеріне әртүрлі дистрофиялар, дисплазиялар дамуы және жасушаның тіршілігін жоғалтуы жатады.
Дистрофиялар – деп жасушаларда зат алмасуларының бұзылыстарынан олардың тіршілігі өзгеруіне әкелетін құрылымдық және функциялық өзгерістерді айтады. Олар мына жолдармен дамуы мүмкін:
● қалыпты жағдайларда жасушаларда кездеспейтін ауытқыған заттардың түзілуімен (мәселен, нәруыздардың күрделі қанттармен кешендері - амилоидтар түзілуі);
● бір қоспалардың басқа қосындыларға артық ауысып кетуімен (мәселен, көмірсуларының майларға, көмірсулары мен майлардың нәруыздарға ауысуы);
● жасуша ішілік құрылымдардың немесе заттардың ыдырап кетуімен (мәселен, мембранадағы нәруыздық-липидтік кешеннің ыдырауы);
● жасушалар мен жасуша аралық заттарда органикалық және бейорганикалық қоспалардың сіңбелері пайда болуымен (мәселен, атеросклероз кезінде артериялардың ішкі қабығы жасушаларында төмен тығыздықты липопротеидтер мен Са2+ -иондарының сіңбелері болуы).
Зат алмасуларының бұзылу түрлеріне қарай нәруыздық (диспротеиноздар), майлық (липидоздар), көмірсулық, пигменттік, минералдық дистрофияларды ажыратады. Олардың жеке тобына жинақталу аурулары жатады.
Дисплазиялар - жасушалардың құрылымы, зат алмасулары мен атқаратын міндеттемелерінің тұрақты өзгерістерімен қабаттасатын олардың дамуындағы нақтыланудың бұзылыстары.Жасушаның нақтылануы оның тектік ақпаратына байланысты.
Жасушалық дисплазия өспе өсуіне тән құбылыс. Бұл кезде жасушалардың көлемі ұлғаяды, пішіні өзгереді, олардың ішінде әртүрлі қоспалар пайда болады. Жасуша ішілік құрылымдардың арақатынасы өзгереді.
Дисплазиялардың бір түрі болып метаплазия есептеледі. Метаплазия - деп белгілі ағза жасушаларының басқа тін жасушаларына айналып кетуін айтады. Мәселен, асқазан жасушалары ішек жасушаларына айналып кетеді.
Жасушалардың тіршілігін жоғалтуы.Жасушалардың тіршігін жоғалтуы некроздық және апоптоздық жолдармен болады.
Некроз (грек. песгоs - өлі) - деп жасуша тіршілігінің қайтымсыз жоғалуын айтадыОл дистрофияның соңғы сатысында немесе төтенше бүліндіргіш ықпалдың тікелей әсерінен дамиды. Тіршілігін жоғалтқан жасушалардың айналасына фагоциттер шоғырланып, қабыну үрдісі байқалады. Некрозға ұшыраған жасушалар лизосомалық ферменттердің және оттегінің белсенді бос радикалдарының қатысуымен ыдыратылады. Жасуша құрылымдарының осындай жолдармен өзін-өзі ыдыратуын аутолиз дейді. Бүлінген және некрозға ұшыраған жасушалардың ыдыратылуы фагоциттердің және микробтардың протеолиздік ферменттерінің қатысуымен болады.
Жасушалардың некрозға ұшырау алдындағы жағдайын некробиоз дейді. Бұл кезде пайда болған өзгерістер қайтымсыз өзгерістерге жатады және некроз дамуына әкеледі.
Апоптоз- жасушаның алдын-ала бағдарланған гендік ақпараты бойынша тіршілігін жоюы. Оның дамуын қадағалайтын екі түрлі гендер белгілі. Апоптоз дамуын тежейтін ген Bcl-2 қадағалауымен антиапоптоздық нәруыз түзіледі. Ол митохондрийлардың мембранасының өткізгіштігін төмендетіп, жасуша цитоплазмасына, апоптозды сергітетін фактор - цитохром С шығарылуын азайтады. Апоптозды күшейтетін нәруыз Bad, Bax гендерімен және супрессор гендер Rb (ретинобластома), p53 гендерімен қадағаланады. Осы нәруыздың түзілуінен эндонуклеаза ферментінің белсенділігі артып, ДНК молекуласы бөлшектеліп кетеді. Апоптоздың гендік бағдарламасын дабылдық ақпарат жүзеге асырады. Ал, некроз бүліндіргіш ықпалдың әсерінен дамиды. Некроздың нәтижесінде жасушаның ыдырауы болып, оның ішіндегі заттар жасуша аралық кеңістікке босап шығады. Босап шыққан лизосомалық ферменттер айналасындағы тіндерді ыдыратады. Ал, апоптоз кезінде жасушаның ыдыраған құрамбөлшектерін фагоциттер жеп қояды. Некроз әрдайым дерттік үрдіс. Ал, апоптоз көптеген табиғи болмыстық үрдістердің барысында байқалады. Сонымен бірге ол бүліндіргіш ықпалға жасушаның бейімделуі түрінде де кездеседі. Некрозға қарағанда апоптоз энергия пайдаланып өтеді және ол нуклеин қышқылдары мен нәруыздар түзілуін қажет етеді.
Апоптоз кезінде жасуша тығыздалып, ядросы ыдырайды, ДНК молекуласы бөлшектенеді. Артынан жасуша бөлшектеніп, апоптоздық денешіктер пайда болады. Оларды макрофагтар мен нейтрофильдер жеп қояды. Бұл кезде ешқашан қабыну дамымайды.
Апоптозды дамытатын дабылдық түрткілер мембрана арқылы әсер ететін немесе жасуша ішілік болып екіге ажыратылады.
Мембрана арқылы әсер ететін дабылдық түрткілерге өспелерді жоятын фактор (ӨЖФ) және CD95L молекуласы жатады. Бұлар, жасуша мембранасындағы сәйкес рецепторларға әсер етіп, апоптоз дамуына әкеледі. Сонымен бірге апоптоз жасушаның бөлініп көбеюін және жетілуін реттейтін өсу факторлары мен цитокиндердің, гормондардың болмауынан да дамуы мүмкін
Жасуша ішілік түрткілерге: жасуша ішінде Н+ -, Са2+ -иондарының жиналып қалуы, бос радикалдар, жасуша ішіндегі вирустар, ядролық рецепторлар арқылы әсер ететін гормондар (глюкокортикоидтық, минералокортикоидтық, тиреоидтық гормондар) жатады. Жасуша ядросында ДНК молекуласының қалпына келуінің бұзылуы да апоптоз дамытады.
Жасуша бүліністерінің түрлері.
Жасушаның бүліністері: жіті және созылмалы, біріншілік (тікелей себепкер ықпалдан) және салдарлық (себепкер ықпалдан пайда болған белсенді биологиялық әсерлі заттардың ықпалдарынан), толық және үлестік (жасуша бөлшектерінің), қайтымды және қайтымсыз, спецификалық және бейспецификалық болып ажыратылады.
Спецификалық бүліністер деп туындатқан себепкер ықпалға ғана тән өзгерістерді ұғады. Бұндай бүліністер сирек кездеседі. Мәселен, осмостық қысымның көтерілуі қандай да болмасын жасушаның ісінуіне, қабығының керілуіне және оның тұтастығының бұзылуына әкеледі. Қанда әлдостеронның деңгейі көтерілуі әртүрлі жасушаларда натрий ионының жиналуына әкеледі. Жарақаттану кезінде жасуша мембранасының жыртылуы, синил қышқылының әсерінен цитохромоксидаза ферментінің белсенділігі жоғалуы осы бүліндіргіш әсерлерге ғана тән өзгерістер.
Көпшілік жағдайларда әртүрлі бүліндіргіш ықпалдар әсер еткеннен кейін жасушаларда біркелкі өзгерістер дамиды. Бұндай өзгерістерді бейспецификалық өзгерістер дейді. Оларға жоғарыда көрсетілген мембрананың өткізгіштігі көтерілуі, рецепторлық, насостық, каналдық қызметтерінің бұзылыстары және мембраналық потенциалдың өзгерістері, жасуша ішінде ацидоз дамуы т.с.с. жатады.
Бүліндіргіш ықпалдарға жасушаның бейімделу жолдары.
Бүліндіргіш әсер әлсіз, немесе жасушаның қалпына келу қабілеті жақсы болғанда, қорғаныстық-икемделістік тетіктер қосылады. Олар әртүрлі бағыттарда болады:
● жасушаның энергиямен қамтамасыз етілуіне қажетті:
♣ жасуша глюкозаны оттегісіз ортада ыдыратып (анаэробтық гликолиз) шамалы болса да энергия (АТФ) түзеді;
♣ тотығу-фосфорлану үрдістеріне қатысатын ферменттердің белсенділігін арттырады;
♣ митохондрийлар ұлғайып, олардың гипертрофиясы дамиды;
♣ жасуша ішінде АТФ энергиясын тасымалдауға қатысатын ферменттер (адениннуклеотидтрансфераза, креатинфосфокиназа) әсерленеді;
♣ АТФ энергиясын пайдалануға қажетті ферменттердің (АТФ-азалардың) белсенділігін күшейтеді;
♣ жасуша өзінің атқаратын қызметін шектеп, энергияға деген мұқтаждығын азайтады;
♣ жасушада, энергия жұмсауды қажет ететін, түзілістік үрдістер азаяды.
● жұқпаларға, химиялық улар мен уыттарға қарсы:
♣ микросомалық ферменттердің белсенділігі артып, уытты заттарды тотықтыру, тотықсыздандыру, метил тобынан айыру т.б. жолдармен уытсыздандырады;
♣ эндоплазмалық торшалардың, рибосомалардың, Голдж кешенінің қызметтері артуы ауру туындататын инфекциялық, химиялық заттарды ыдырататын ферменттердің, нәруыздардың, гормондардың, вирустарға қарсы интерферондардың түзілуін күшейтеді;
♣ жасуша буферлік жүйелерін қосып, қышқылдық-сілтілік үйлесімді реттейді.
● жасуша мембранасы бүліністеріне қарсы:
♣ антиоксиданттық ферменттердің ( супероксиддисмутаза, каталаза, глютатионпероксидаза т.б.) түзілуін арттырады;
♣ бүлінген мембрана бөлшектерін қалпына келтіреді.
● ДНК молекуласының гендік ақаулары кездерінде:
♣ бүлінген немесе бөтен текті бөлшегін дер кезінде тауып аластайды;
♣ оның үзілген жерлерін жояды;
♣ оның бүлінген бөлшегін қалыпты бөлшектермен ауыстырады;
♣ онда транскрипция, трансляция үрдістерін қалпына келтіреді.
Артынан жасушаның және оның кейбір бөлшектерінің қалпына келуі (регенерациясы), гипертрофиясы мен гиперплазиясыдамиды.
Осы келтірілгендермен бірге бүгінгі күні жасушаға бүліндіргіш ықпал
(температураның өзгерістері, гипокселтірілгендермен бірге бүгінгі күні жасушаға бүліндіргіш ықпал
(температураның өзгерістері, гипоксия, химиялық заттар, вирустық жұқпа т.с.с.) әсер еткенде арнайы бір нәруыздар өндірілетіні белгілі болды. Ол нәруыздарды ағылшынша - heat shock proteins (HSP), орысша - белки теплового шока немесе белки стресса дейді, ал қазақша оларды ауырпалық нәруыздары деуге болады. Бұл нәруыздар жасушаны бүліндіргіш ықпалдардан қорғап, оның тіршілігін сақтап қалуға көмектеседі. Олардың ішінде молекулалық массасы 70 000 (hsp 70), 90 000 (hsp 90) ауыртпалық нәруыздары кең тараған. Бұлар басқа нәруыздардың жиналуын, жиырылып жазылуын реттеуге қатысады. Осыдан жасушада нәруыздық топтамалардың жиналып қалуынан сақтандырады, жасушаның төзімділігін көтереді.
Жасушаның бүлінген бөлшектері фагоциттерді өзіне тартатын заттар шығарады. Фагоциттер бүлінген бөлшектерді жеп қояды. Жасуша ішіндегі құрылымдардың гиперплазиясы дамып, жойылған жасуша бөлшектерінің орны толтырылады. Гиперплазия молекулалардың деңгейінде (гендердің саны артуы, жаңа молекулалардың түзілуі күшеюі), жасуша ішілік құрылымдарда (митохондрийларда, рибосомаларда, лизосомаларда ж. б.) және жасушалық деңгейлерде байқалады. Көрсетілген икемделістік тетіктер жеткіліксіз болғанда жасушаның тіршілігі қайтымсыз жоғалады.
ЖАСУША БҮЛІНІСТЕРІН ЕМДЕУ ЖОЛДАРЫНЫҢ НЕГІЗДЕРІ.
Жасуша бүліністерін емдеу ең алдымен шақырған себепкер ықпалды аластауға бағытталуы қажет. Сонымен бірге бүлінген жасушалардың мембраналарын тұрақтандыратын дәрі-дәрмектер пайдаланылады. Мембраналардың бүліністерінің негізінде еркін радикалды тотығу мен май қышқылдарының асқын тотығуы болғандықтан оларды ескертетін табиғи және жасанды антиоксиданттар қолданылады. Мембраналарда фосфолипаза, липаза, протеаза ферменттерінің белсенділігі көтерілуіне қарсы олардың бәсеңсіткіштері пайдаланылады. Жасуша ішіне Са2+ иондары көп енуінен сақтандыру үшін кальцийдің шабан өзекшелерінің тежегіштері қолданылуы қажет. Сонымен бірге мембраналарды тұрақтандыратын дәрі-дәрмектер (глюкокортикоидтық, анаболизмдік гормондар), адреноблокаторлар пайдаланылады.