Иммобилизованные ферменты
(от лат. immobiiis - неподвижный), препараты ферментов, молекулы которых связаны с матрицей, или носителем (как правило, полимером), сохраняя при этом полностью или частично свои каталитич. свойства. иммобилизованные ферменты обычно не раств. в воде. между двумя фазами возможен обмен молекулами субстрата, продуктов каталитич. реакции, ингибиторов и активаторов. Существует неск. осн. способов иммобилизации ферментов: 1) путем образования ковалентных связей между ферментом и матрицей; 2) полимеризацией мономера. образующего матрицу, в присутствии фермента, который при этом оказывается включенным в сетку полимера - обычногеля. 3) благодаря электростатич. взаимод. противоположно заряженных групп фермента и матрицы; 4)сополимеризацией фермента и мономера. образующего матрицу; 5) связыванием фермента и матрицы в результате невалентных взаимод. - гидрофобных, с образованием водородных связей и др.; 6) инкапсулированием - созданием около молекул фермента полупроницаемой капсулы, например, включением фермента в липосомы. 7) сшиванием молекул фермента между собой, например, глутаровым альдегидом. диметиловым эфиром диимида адипиновой кислоты. Особый случай иммобилизации проведение ферментативных реакций в двухфазной системе, когда фермент находится в водной фазе, а субстраты и продукты реакции распределяются между орг. и водной фазами, что позволяет в зависимости от коэф. распределения веществ между фазами сдвигать равновесие реакции в нужную сторону;диспергирование фаз увеличивает пов-сть их раздела и тем самым улучшает доступ субстрата к ферменту. Среди способов иммобилизации наиб. распространение получили ковалентное связывание фермента с матрицей и включение фермента в гель. В первом случае в качестве матрицы обычно используют целлюлозу, декстрановые гели (сефароэу, агарозу), микропористые стекла или кремнеземы, а также синтетич. полимеры. Матрицу при ковалентной иммобилизации ферментов обычно предварительно активируют, обрабатывая, например, бромцианом, азотистой кислотой или цианурхлоридом. Благодаря этому она становится носителем активных группировок, которые способны вступать в реакцию сочетания, взаимод. с группами NH2, ОН, СООН. Во втором случае в качестве гелеобразующегополимера используют полиакриламид. На практике иммобилизация часто осуществляется одновременно неск. способами. Так, при фиксации ферментов ковалентными связями между их молекулами и матрицей обычно возникают также нековалентные взаимодействия. Известны способы предварит. хим. модификации молекул фермента низкомол. веществами или растворимыми полимерами, имеющими заряженные группировки, что изменяет у таких модифицир. белков электростатич. заряд молекулы и позволяет достаточно прочно сорбировать их на ионообменных смолах. При всех типах иммобилизации матрица, взаимодействуя с ферментом, может инактивировать последний или создавать пространств. затруднения для доступа субстрата к активному центру. При ковалентном связывании фермента для предотвращения отрицат. влияния матрицы между ней и молекулой фермента вводят разобщающую цепь атомов - спейсер (наз. также "вставкой" или "ножкой"). Кроме того, часто стремятся использовать для иммобилизации гидрофильные матрицы, создающие вблизи фермента более естеств. микроокружение. При иммобилизации ферментовнеобходимо, чтобы активные группы матрицы не блокировали каталитич. центр фермента, а условия иммобилизации не приводили к потере его активности. Определенные ограничения на способ иммобилизации налагают и особенности
20.смолами и обесцвечивают активированным углем. Далее сироп уваривают при 60 ºС (при более высоких температурах фруктоза рагзлагается) в выпарных аппаратах пленочного типа до содержания СВ 71…74 %, охлаждают до 30 ºС и хранят при 25…30 ºС, так как при температуре ниже 25 ºС начинается кристаллизация глюкозы, а при температуре выше 30 ºС нарастает цветность сиропа из-за разложения моносахаридов.
Глюкозно-фруктозные сиропы находят широкое применение при производстве детского и диетического питания, хлебобулочных изделий, кремов, тортов, пирожных, джемов, помадки, зефира, пастилы.
По своим свойствам такие сиропы близки к инвертному. Из-за большого содержания моносахаридов, особенно фруктозы, использование сиропов позволяет получать кондитерские изделия повышенного качества: они долго остаются свежими и не засыхают. Хлебобулочные изделия, приготовленные на глюкозно-фруктозном сиропе, имеют лучшую окраску корки и более длительный срок сохранения свежести. Сироп с содержанием фруктозы 90 % позволяет получать пищевые продукты пониженной энергетической ценности благодаря снижению содержания сахара в рецептуре изделий за счет более сладкого вкуса сиропа.
2 АКТИВНЫЙ ЦЕНТР ФЕРМЕНТОВ
Каталитическая функция ферментов определяется наличием одного или нескольких активных центров.
Активный центр – это участок в пространственного структуре фермента, с которым связывается субстрат и подвергается химическому превращению. Число активных центров может быть равно числу субъединиц в четвертичной структуре фермента, т.е. сколько субъединиц (протомеров), столько активных центров.
В активном центре условно выделяют два участка:
- контактный (якорный или субстратный), отвечающий за специфичность связывания субстрата (узнавание);
- каталитический, где происходит химическое превращение субстрата после его связывание (сначала фермент узнает субстрат, притягивает его, затем субстрат располагается в этом активном центре.
Структурная организация фермента
1. Особенности образования активного центра у ферментов протеинов (простых белковых ферментов).
Обычно он образован 12-16 аминокислотными остатками полипептидной цепи. Иногда их число больше. Аминокислоты, формирующие активный центр, находятся в разных местах полипептидной цепи. При пространственной укладки белка-фермента (в третичную структуру), они сближаются и образуют активный центра.
Приблизительно 1/2 – 1/3 аминокислот фермента прямо или косвенно участвуют в работе активного центра.
2.Особенности образования активного центра у ферментов-протеидов (сложных белков-ферментов).
Протеиды состоят из:
Апофермент (белковая часть) + кофактор (небелковая часть) = холофермент (активный комплекс).
Кофактор (или простетическая группа) чаще всего предствавлен витаминами или ионами металлов.
Холофермент в диссоциированном состоянии неактивен.
При низкой температуре происходит обратимая инактивация фермента, т.к. наблюдаются незначительные изменения конформации активного центра фермента.
Фермент имеет белковую природу, поэтому температура на него, влияет также как на белок (повышении температуры приводит к денатурации).
3 НОМЕНКЛАТУРА И КЛАССИФИКАЦИЯ ФЕРМЕНТОВ
Каждый фермент имеет 2 названия. Первое - короткое, так называемое рабочее, удобное для повседневного использования. Второе (более полное) - систематическое, применяемое для однозначной идентификации фермента.
А. Рабочее названиеВ названии большинства ферментов содержится суффикс "аза", присоединённый к названию субстрата реакции, например уреаза, сахараза, липаза, нуклеаза или к названию химического превращения определённого субстрата, например лактатдегидрогеназа, аденилатциклаза, фосфо-глюкомутаза, пируваткарбоксилаза.
8 ВЛИЯНИЕ ТЕМПЕРАТУРЫ НА АКТИВНОСТЬ ФЕРМЕНТОВ
Ферменты обладают всеми свойствами белков. Однако по сравнению с белками, выполняющими другие функции в клетке, ферменты имеют ряд специфических, присущих только им свойств.
Зависимость активности ферментов от температуры.Температура может влиять по-разному на активность фермента. При высоких значениях температуры может происходить денатурация белковой части фермента, что негативно сказывается на его активности. При определенных (оптимальных) значениях температура может влиять на скорость образования фермент-субстратного комплекса, вызывая увеличение скорости реакции. Температура, при которой каталитическая активность фермента максимальна, называется температурным оптимумомфермента. Различные клеточные ферменты имеют собственные температурные оптимумы, которые определяются экспериментально. Для ферментов животного происхождения температурный оптимум находится в интервале 40 - 50°С
Установлено, что скорость ферментативных реакций при изменении температуры инкубации на 10 °С изменяется в 2 раза. Например, активность АСТ в сыворотке фирмы Randox, определённая при 37 °С, составляет 35 U/л, а при 25 °C – 16 U/л. При дальнейшем понижении температуры реакционной смеси скорость реакции будет снижаться: при 15 °С активность АСТ равна 8 U/л, при 5 °С — 4 U/л. Поэтому определение активности ферментов необходимо всегда проводить при температуре, указанной в инструкции по использованию набора.
Температура, при которой наблюдается максимальная активность ферментов, называется оптимальной.Для большинства ферментовоптимальной температурой является температура от +35С — +45С. Если фермент поместить в условия, ниже оптимальной температуры, будет происходить снижение его активности, такое состояние называется обратимой инактивацией фермента, т.к. если вновь поднять температуру до оптимальной, активность фермента возобновится. Если поместить фермент в условия, где температура будет выше оптимальной, то также будет прроисходить снижение его активности, но в данном случае необратимая инактивация,т.к. если понизить температуру до оптимальной, активность фермента не возобновится. Это объясняется тем, что высокая температура вызывает денатурацию молекулы фермента.
13. ТЕХНОЛОГИЯ КУЛЬТИВИРОВАНИЯ МИКРООРГАНИЗМОВ
Производство ферментных препаратов микробного происхождения может осуществляться поверхностным и глубинным методами.
Поверхностный метод заключается в культивировании микроорганизмов на поверхности увлажненной стерилизованной сыпучей питательной среды, размещенной в кюветах. Инкубацию микроорганизмов ведут в специальном термостатируемом цехе при постоянном контроле в нем температуры, влажности и расхода воздуха. Основные параметры поверхностного способа получения ферментов приведены на слайде.
Для выращивания продуцентов ферментов глубинным методом в промышленных условиях используют ферментаторы из нержавеющей стали, снабженные устройствами для перемешивания и подачи в жидкую питательную среду стерильного воздуха.
Глубинный способ ведения процесса имеет ряд существенных преимуществ перед поверхностным культивированием, т. к. позволяет существенно автоматизировать процесс, в ряде случаев значительно сократить объёмы отходов, проводить процесс непрерывно, сократить в 2 – 4 раза площади цехов, а также позволяет использовать анаэробных продуцентов.
Сначала ферментатор заполняют питательной средой, стерилизуют ее, затем засевают чистой культурой, подаваемой из специального генератора. Для предотвращения инфицирования в ферментере поддерживают повышенное давление наряду с оптимальными значениями рН, температуры, окислительно-восстановительного потенциала и другими условиями культивирования. Основные параметры глубинного способа получения ферментов приведены на слайде.
Культура микроорганизмов, выращенная поверхностным способом, и культуральная жидкость после глубинного культивирования содержат большое количество балластных веществ. Выделение и очистка ферментов – трудоёмкий и дорогостоящий процесс поэтому, если ферментный препарат можно использовать в виде неочищенной культуры микроорганизмов, его очистку не проводят. В таких отраслях, как спиртовая и кожевенная, целесообразнее использовать именно неочищенную культуру микроорганизма; то же самое можно сказать и об использовании культур микроорганизмов в сельском хозяйстве при приготовлении комбикормов и при непосредственной обработке кормов на фермах.
В большинстве отраслей пищевой промышленности (хлебопекарной, пивоварении, виноделии, сыроделии, крахмало-паточном и сокоэкстрактном производствах), а также в микробиологической промышленности и особенно медицине можно использовать только очищенные препараты ферментов, частично или полностью освобожденные от балластных веществ.
Исходным материалом для получения очищенных ферментных препаратов может служить фильтрат культуральной жидкости, реже – биомасса продуцента или водный экстракт из поверхностной культуры продуцента. Ферментные препараты могут быть получены в виде порошков или жидких концентратов. В процессе выделения происходит повышение доли активного белка в общей массе препарата, т. е. увеличивается его удельная активность.
Из нее ясно, что экстракт из поверхностной культуры или фильтрат культуральной жидкости является исходным материалом для получения препаратов
субстрата. Так, в случае высокомол. субстратов нельзя использовать методы инкапсулирования или включения фермента в гель. Если матрица несет на себе заряды, то заряд субстрата влияет на кинетич. параметры реакции: разноименные заряды на носителе и субстрате увеличивают скорость реакции, катализируемой иммобилизованные ферменты, одноименные заряды ее снижают и м. б. причиной полной потери активности препарата. Заряды носителя и субстрата влияют также на величину рН, при которой скорость ферментативной реакции максимальна. Важную роль играет распределение субстрата между фазами иммобилизованного фермента и раствора. Ограниченная доступность субстрата к активному центру фермента может привести к изменению специфичности последнего. Особенно это Характерно для высокомол. субстратов, которые из-за малого коэф. диффузии медленно переходят в фазу иммобилизованные ферменты, что приводит к относит. увеличению скоростей др. реакций с участием субстратов меньших размеров. В некоторых случаях возможно также изменение направления реакции. Так, фермент эндополигалактуроназа, катализирующий расщепление полигалактуроновой кислоты в середине молекулы, после иммобилизации отщепляет низкомол. фрагменты от концов молекулы. Существ. влияние на кинетику реакций, катализируемых иммобилизованными ферментами, оказывают два диффузионных барьера - внешний и внутренний.
16. Физические методы иммобилизации
Нерастворимые носители
Фермент можно адсорбировать на твердом носителе за счет электростатических, гидрофобных или водородных связей. Именно с физической адсорбции началась в 1916 г. история иммобилизованных ферментов. Этот метод до сих пор используется во многих отраслях промышленности.
Удастся ли белку прочно прикрепиться к носителю, зависит от двух особенностей этого носителя: удельной поверхности и количества пор. Чем больше удельная поверхность, тем больше «посадочных мест» для белка; чем лучше развиты поры, тем ему легче закрепиться.
Водный раствор фермента просто помещают на подложку, либо смешивают с частицами носителя, или пропускают через заполненные носителем колонки. Например, осадок гидроксида титана заливают раствором фермента, перемешивают и сушат. При этом каждый грамм осадка впитывает не менее 64 мг белка.
Физическая адсорбция хороша тем, что полностью сохраняет активность фермента – ведь молекула его не образует новых химических связей, а значит не изменяется. Оборотной стороной этого достоинства является невысокая прочность прикрепления белка. Он сравнительно легко отрывается от поверхности, что приводит не только к потере самого фермента, но и к загрязнению продуктов реакции.
Прочность повышают, заранее обрабатывая носитель ионами металлов, полимерами, гидрофобными соединениями, покрывая его слоем белка или липида. Можно обработать и сам фермент, но это снижает его активность.
Носители:кремнезем, активированный уголь, графитовая сажа, различные глины, пористое стекло, полисахариды, синтетические полимеры, оксиды алюминия, титана и других металлов, целлюлоза, декстрановые гели (сефароза, агароза).
Гели
Самый простой способ иммобилизовать фермент – включить его в трехмерную матрицу геля. Этот способ подходит не только для отдельных ферментов, но и для их комплексов и даже целых клеток.
22. Классификация носителей для ферментов
Для получения иммобилизованных ферментов используется ограниченное число как органических, так и неорганических носителей. К носителям предъявляются следующие требования (Дж.Порат, 1974):-
-высокая химическая и биологическая стойкость; -
-высокая химическая прочность;
-достаточная проницаемость для фермента и субстратов, пористость, большая удельная поверхность;
-возможность получения в виде удобных в технологическом отношении форм (гранул, мембран);
-легкая активация;
-высокая гидрофильность;
-невысокая стоимость.
Классификация носителей 1)органические (низкомолекулярные и полимерные) 2)Не органические (макропористые и другие)
Следует отметить, что органические носители (как низко-, так и высокомолекулярные) могут быть природного или синтетического происхождения. Природные полимерные органические носители делят в соответствии с их биохимической классификацией на 3 группы: полисахаридные, белковые и липидные.
Синтетические полимеры также можно разделить на группы в связи с химическим строением основной цепи макромолекул: полиметиленовые, полиамидные, полиэфирные.
Для иммобилизации ферментов наиболее широко используются природные полисахариды и синтетические носители полиметильного типа, остальные применяются значительно реже. Большое значение природных полимеров в качестве носителей для иммобилизации объясняется их доступностью и наличием реакционно-способных функциональных групп, легко вступающих в химические реакции. Характерной особенностью этой группы носителей также является их высокая гидрофильность. Недостаток природных полимеров - неустойчивость к воздействию микроорганизмов и довольно высокая стоимость.
Наиболее часто для иммобилизации используются такие полисахариды, как целлюлоза, декстран, агароза и их производные. Целлюлоза гидрофильна, имеет много гидроксильных групп, что позволяет модифицировать её, замещая эти группы. Для увеличения механической прочности целлюлозу гранулируют путем частичного гидролиза, в результате которого разрушаются аморфные участки. На их место для сохранения пористости между кристаллическими участками вводят химические сшивки. Гранулированную целлюлозу довольно легко превратить в различные ионообменные производные, такие как ДЭАЭ-целлюлоза, КМЦ и т.д.
Широко распространены носители на основе декстрана, выпускаемые под названием "сефадексы". При высушивании они легко сжимаются, в водном растворе сильно набухают. В этих носителях размер пор в геле регулируется степенью сшитости. К группе декстранов относят и крахмал. Химически модифицированный крахмал сшивается агентами, такими как формальдегид. Таким способом был получен губчатый крахмал, обладающий повышенной устойчивостью по отношению к ферментам, гидролизу. Водорастворимые препараты на основе декстрана часто применяются как носители лекарственных средств в медицине.
Хорошим носителем считается агар. Его свойства улучшаются после химической сшивки, например, диэпоксидными соединениями. Такой агар становится устойчивым к нагреванию, прочен, легко модифицируется.
| Б. Классы ферментов
Международный союз биохимии и молекулярной биологии в 1961 г. разработал систематическую номенклатуру, согласно которой все ферменты разбиты на 6 основных классов в зависимости от типа катализируемой химической реакции. Каждый класс состоит из многочисленных подклассов и подподклассов с учётом преобразуемой химической группы субстрата, донора и акцептора преобразуемых группировок, наличия дополнительных молекул и т.д. Каждый из 6 классов имеет свой порядковый номер, строго закреплённый за ним.
Классификация ферментов.В настоящее время известно более 2000 ферментов. Все ферменты разделены на шесть классов, каждый из которых имеет строго определенный номер. Оксидоредуктазыкатализируют окислительно-восстановительные процессы.
1. Трансферазыкатализируют реакции переноса функциональных групп и молекулярных остатков с одной молекулы на другую.
2. Гидролазыкатализируют реакции гидролиза.
3. Лиазыкатализируют реакции отщепления (кроме атомов водорода) с образованием двойной связи либо присоединения по двойной связи, а также негидролитический распад органических соединений либо синтез без участия макроэргических веществ.
4. Изомеразыкатализируют процессы изменения геометрической или пространственной конфигурации молекул. Лигазыкатализируют реакции синтеза, сопровождающиеся гидролизом богатой энергией связи (как правило, АТФ
4 ХАРАКТЕРИСТИКА КЛАССА ОКСИДОРЕДУКТАЗ. ПРИМЕРЫ
Ферменты этого класса катализируют окислительно-восстановительные реакции, лежащие в основе биологического окисления. Класс насчитывает 22 подкласса. Коферментами этого класса являются НАД,НАДФ, ФАД, ФМН, убихинон, глутатион, липоевая кислота.
Примером подклассов могут служить ферменты, действующие на СН-ОН-группу доноров, на СH-СН-группу доноров, на СН-NН2-группу доноров, на гемсодержащие доноры.
Наиболее распространены следующие рабочие названияоксидоредуктаз:
1. Дегидрогеназы – оксидоредуктазы, катализирующие дегидрирование субстрата с использованием в качестве акцептора водорода любых молекул, кроме кислорода.
2. Если перенос водорода от молекулы донора трудно доказуем, то такие оксидоредуктазы называют редуктазами.
3. Оксидазы – оксидоредуктазы, катализирующие окисление субстратов с молекулярным кислородом в качестве акцептора электронов без включения кислорода в молекулу субстрата.
4. Монооксигеназы – оксидоредуктазы, катализирующие внедрение одного атома кислорода в молекулу субстрата с молекулярным кислородом в качестве донора кислорода.
5. Диоксигеназы – оксидоредуктазы, катализирующие внедрение 2 атомов кислорода в молекулу субстрата с молекулярным кислородом в качестве донора кислорода.
6. Пероксидазы – оксидоредуктазы, катализирующие реакции с пероксидом водорода в качестве акцептора
9 ВЛИЯНИЕ рН СРЕДЫ НА АКТИВНОСТЬ ФЕРМЕНТОВ
.Большинство ферментов проявляет максимальную активность при значениях рН, близких к нейтральным. Лишь отдельные ферменты "работают" в сильно кислой или сильно щелочной среде. Например, активность пепсина - фермента, гидролизующего белки в желудке, - максимальна при рН 1,5 - 2,5. В щелочной среде "работают" ферменты, локализованные в кишечнике. Изменение оптимального для данного фермента значения рН-среды может привести к изменению третичной стурктуры фермента, что скажется на его активности. С другой стороны, при изменении рН может измениться ионизация субстрата, что повлияет на образование фермент-субстратного комплекса.
рН среды влияет на заряд молекулы фермента, а значит на работу АЦ. Оптимальная рН для каждого фермента своя, но для большинства ферментов от 4 до 7. Например, для альфа-амилазы слюны опт.рН равна 6,8.
Зависимостьактивности ферментовот рН среды.Ферменты обычно наиболее активны в пределах узкой зоны концентрации водородных ионов, соответствующей для животных тканей в основном выработанным в процессе эволюции физиологическим значениям рН среды 6,0–8,0.. При определении зависимостиактивности фермента от концентрации водородных ионов реакцию проводят при разных значениях рН среды, обычно при оптимальной температуре и наличии достаточно высоких (насыщающих) концентраций субстрата.
рН-оптимум действия ферментов лежит в пределах физиологических значений. Исключение составляют пепсин, рН-оптимум которого 2,0 (при рН 6,0 он не активен и не стабилен). Объясняется это, во-первых, структурной организацией молекулы фермента и, во-вторых, тем, что пепсин является компонентом желудочного сока, содержащего свободную соляную кислоту, которая создает оптимальную кислую среду для действия этого фермента. С другой стороны, рН-оптимум аргиназы лежит в сильнощелочной зоне (около 10,0); такой среды нет в клетках печени, следовательно, in vivo аргиназа функционирует, по-видимому, не в своей оптимальной зоне рН среды.
Согласно современным представлениям, влияние изменений рН среды на молекулу фермента заключается в воздействии на состояние и степень ионизации кислотных и основных групп (в частности, СООН-группы дикар-боновых аминокислот, SH-группы цистеина, имидазольного азота гисти-дина, NH2-группы лизина и др.). При резких сдвигах от оптимума рН среды ферменты могут подвергаться конформационным изменениям, приводящим к потереактивности вследствие денатурации или изменения заряда молекулы фермента. При разных значениях рН средыактивный центр может находиться в частично ионизированной или неионизированной форме, что сказывается натретичной структуре белка и соответственно на формировании активного фермент-субстратного комплекса. Имеет значение, кроме того, состояние ионизации субстратов и кофакторов
ферментов различной степени очистки. На первом этапе выделения отходом процесса является нерастворимая часть культуры – биошрот, содержащий нерастворимые включения среды и биомассу продуцента.
Далее в зависимости от свойств выделяемого фермента и сопутствующих ему балластных веществ при получении очищенных ферментных препаратов комбинируют различные приемы и методы, такие, как термическое фракционирование,осаждение органическими растворителями и солями, очистка на молекулярных ситах, ионообменная хроматография, электрофорез и др.
Еще одна технологическая схема получения препаратов из поверхностной и глубинной культур в виде жидких концентратов, сухих технических препаратов, получаемых сушкой распылением, и препаратов, осажденных органическими растворителями.
Фильтрат охлажденной культуральной жидкости собирается в основном сборнике и по мере надобности передается в сборник небольшой вместимости перед поступлением в подогреватель вакуум-выпарной установки пленочного типа. Концентрат культуральной жидкости с содержанием сухого вещества 6 – 10 % поступает в сборник концентрата. Для получения сухого технического препарата концентрат направляют в башню распылительной сушилки 8. Сухой препарат через циклон 10, бункер 11 и шнек 12 попадает на стадию стандартизации, фасования и упаковывания.
Для получения более очищенного препарата концентрат из сборника подается на осаждение органическим растворителем. Предварительно концентрат охлаждают в теплообменнике до температуры 2 – 3 °С и подают через дозатор в осадитель. Одновременно в осадитель дозируется охлажденный растворитель. Образовавшийся осадок отделяют на сепараторе 16. Надосадочную жидкость направляют на регенерацию, а осадок – на промывку спиртом и повторное сепарирование. Промытый осадок высушивают в вакууме, измельчают, взвешивают, смешивают с наполнителем и направляют на фасование и упаковывание.
При получении ферментных препаратов из культур микроорганизмов, выращенных поверхностным способом, процесс очистки начинается с экстракции ферментов водой. Нерастворимый осадок высушивают и в виде сухого биошрота утилизируют на корм скоту.
Экстракт с содержанием сухого вещества 7 – 14 % при получении из него сухих препаратов не нуждается в дополнительном концентрировании и поэтому может быть сразу направлен на распылительную сушку с целью получения технического препарата, или же экстракт направляется в охладитель, а затем на осаждение органическими растворителями или солевыми растворами. Из экстракта можно получать стабильный жидкий концентрат с содержанием сухого вещества 50%, для чего экстракт направляют в сборник, затем в подогреватель и на вакуум-выпарную установку. Готовый жидкий концентрат фасуют в специальные емкости и направляют на склад готовой продукции. Из глубинной культуры можно также получать жидкие концентраты, например, методом ультрафильтрации.
Есть два способа включения фермента в гель. Чтобы получить гель, нужно сначала соединить отдельные молекулы в полимерные цепочки, а затем сплести из этих цепочек трехмерную структуру. Таким образом, процесс имеет две стадии, и на каждой из них может быть добавлен фермент. Можно ввести его в водный раствор будущего носителя, а затем провести их совместную полимеризацию. При этом молекулы белка сами собой окажутся в толще геля. Другой подход – введение фермента в уже готовый полимер. Затем из него получают гель, несущий в себе фермент.
Преимущество иммобилизации в геле – равномерное распределение фермента во всем объеме носителя. Кроме того, большинство гелей устойчиво к механическому, химическому, тепловому воздействию. Включенный в толщу геля фермент можно использовать многократно. Естественное ограничение метода – он пригоден только для растворимых в воде субстратов.
Носители: совместная полимеризация с носителем - полиакриламид, поливиниловый спирт, поливинил-пирролидон, силикагель; введение в готовый гель - крахмал, агар-агар, каррагинан, агароза, фосфат кальция.
Мембраны.Фермент можно отделить от субстрата полупроницаемой перегородкой – мембраной. Мелкие молекулы субстрата будут свободно проходить через нее, а крупные молекулы фермента останутся внутри. Мембрана, в зависимости от состава, может называться микрокапсулой или липосомой. Чем отличаются липосомы от микрокапсул? Липосома – это круглая или удлиненная оболочка, образованная двойным слоем липидных молекул (обычно лецитина). Раствор лецитина в органическом растворителе упаривают. Получается тонкая пленка лецитина, которую рассеивают в виде мелких частиц в водном растворе фермента. Эти частицы сами собираются в двуслойные оболочки вокруг молекул фермента.
17. Химические способы иммобилизаци
Химическая иммобилизация – это создание ковалентных связей между ферментом и носителем. В чем преимущества полученных этого способа? Первое из них – прочность. Ковалентная связь надежно и необратимо прикрепляет фермент к носителю. Какими бы ни были условия реакции – кислотность среды и температура, фермент не отрывается от подложки и не загрязняет целевые продукты. Такое загрязнение недопустимо в производстве лекарств, переработке пищевого сырья, а также при проведении исследований. Второе преимущество – образуя химические связи, можно направленно менять свойства белка. Например, повысить каталитическую активность фермента, стабильность или субстратную специфичность.
Белок взаимодействует с носителем своими аминными, гидроксильными и карбоксильными группами. Носитель для химической иммобилизации необходимо предварительно активировать – бромцианом, азотистой кислотой или цианурхлоридом. После такой обработки на нем появляются функциональные группы, способные захватывать белок.Существуют разные способы химическом иммобилизации, в зависимости от того, за какие группы на поверхности носителя цепляется белок.
При всех достоинствах химической иммобилизации, в промышленности она малоприменима из-за сложности и дороговизны. Этот способ больше подходит для научных исследований.
Аминогруппы. В данном случае используются первичные аминогруппы носителя, связанные с ароматическим кольцом. Превращенные в соли диазония, они взаимодействуют с множеством радикалов. Это могут бытьфенольные, имидазольные, аминные, гуанидиновые, тиольные группы. Например, фенольные радикалы тирозина в щелочной среде образуют азо-соединения. Этот метод привлекателен тем, что аминогруппы могут быть введены в самые разные носители.
Производные карбоксильной группы. Если на поверхности носителя много ацильных групп, к ним можно прикрепить аминогруппы белка. В этом могут помочь ангидриды, галогенангидриды, активированные эфиры и другие производные карбоновых кислот. Лучше всего соединяются с белком галогенангидриды, хуже – эфиры.
Белки в качестве носителей обладают рядом достоинств: вместительны, способны к биодеградации, могут применяться в качестве тонкой (толщиной 80 мкм) мембраны. Иммобилизацию ферментов на белковых носителях можно проводить как в отсутствие, так и в присутствии сшивающих агентов. Белки используются и в фундаментальных биологических исследованиях, и в медицине. К недостаткам белков в качестве носителей относят их высокую иммуногенность (за исключением коллагена и фибрина). Наиболее для иммобилизации используются структурные (кератин, фибрин, коллаген), двигательные (миозин) и транспортные (альбумин) белки.
Синтетические полимерные носители применяются для ковалентной и сорбционной иммобилизации ферментов, для получения гелей, микрокапсул. Полимеры на основе стирола применяются сорбционной иммобилизации. Они могут иметь макропористую, изопористую структуру, а также гетеропористую структуру. Для получения полимерных гидрофильных носителей широко используется акриламид - производное акриловой кислоты.
Широкое распространение получил метод включения ферментов и клеток в полиакриламидный гель, имеющий жесткую пространственную сетчатую структуру. Полиакриламидный гель устойчив к химическим воздействиям. Очень интересную группу представляют полиамидные носители. Это группы различных гетероцепных полимеров с повторяющейся амидной группой -С(О)-NH-. Например, полимеры на основе N-винилпирролидона используются для получения иммобилизованных ферментов, способных медленно распадаться в организме. Кроме того, они биологически инертны, что особенно важно при использовании в медицинских целях. Существенным недостатком большинства полимерных носителей является их способность накапливаться в организме. В этом отношении предпочтение отдается природным полимерам, которые гидролизуются ферментами. Поэтому в состав лекарственных препаратов часто входит декстран, а из синтетических носителей - полимеры на основе N-винилпирролидона. В настоящее время ведутся эксперименты по созданию синтетических полимеров, расщепляющихся с образованием нетоксичных продуктов обмена.
| электронов.
ДЕГИДРОГЕНАЗЫ. Это ферменты, которые катализируют отщепление или присоединение водорода. Дегидрогеназы бывают разные, различают несколько подклассов ДГ.
Например,алькогольДГ является анаэробной, т. к. эта реакция протекает без
участия О2
ОКСИДАЗЫ. Они удаляют водород из субстрата. Они также используют кислород, но в отличие от аэробных дегидрогеназ, их продуктом является молекула воды. Эти ферменты обязательно содержат медь или железо, т.е. металлы с переменными валентностями. Эти ферменты используют в качестве субстрата перекись водородаили органические перекиси, например перекиси липидов. Т.е. это как раз те ферментные системы, которые обезвреживают Н2О2 и другие перекиси.
5 МЕХАНИЗМ ФЕРМЕНТАТИВНОГО КАТАЛИЗА
Ферментативный катализ (биокатализ), ускорение биохимических реакций при участии белковых макромолекул. называемых ферментами (энзимами). Ферментативный катализ - разновидность катализа, хотя термин "ферментация" (брожение)известен с давних времен, когда еще не было понятия химического катализа.
Важнейшие особенности ферментативного катализа - эффективность, специфичность и чувствительность к регуляторным воздействиям. Ферменты увеличивают скорость химического превращения субстрата по сравнению с неферментативной реакцией в 109-1012 раз. Столь высокая эффективность обусловлена особенностями строения активного центра. Принято считать, что активный центр комплементарен переходному состоянию субстрата при превращении его в продукт. Благодаря этому стабилизируется переходное состояние и понижается активационный барьер реакции. Большинство ферментов обладает высокой субстратной специфичностью, то есть способностью катализировать превращение только одного или нескольких близких по структуре веществ. Специфичность определяется топографией связывающего субстрат участка активного центра. Активность ферментов регулируется в процессе их биосинтеза (в том числе благодаря образованию изоферментов, которые катализируют идентичные реакции, но отличаются строением и каталитическими свойствами), а также условиями среды (рН, температура, ионная сила раствора) и многочисленными ингибиторами и активаторами, присутствующими в организме. Ингибиторами и активаторами могут служить сами субстраты (в определенных концентрациях), продукты реакции, а также конечные продукты в цепи последовательных превращений вещества (см.Регуляторы ферментов).Ферментативные реакции чувствительны к внешним условиям, в частности к ионной силе раствора и рН среды. Основные направления современных исследований ферментативного катализа - выяснение механизма, обусловливающего высокие скорости процессов, высокую селективность (специфичность действия ферментов), изучение механизмов контроля и регуляции активности ферментов. Оказалось, в частности, что реакции ферментативного катализа включают большое число стадий с участием лабильных промежуточных соединений, времена жизни которых изменяются в нано- и миллисекундном диапазонах. На активных центрах ферментов протекают быстрые (нелимитирующие) стадии, в результате чего понижается энергетический барьер для наиболее трудной, лимитирующей стадии.
10 АКТИВАТОРЫ И ИНГИБИТОРЫ ФЕРМЕНТОВ
Вещества, которые оказывают влияние на активность ферментов, называют эффекторами. Это могут быть ингибиторы – соединения, тормозящие каталитический процесс, илиактиваторы – вещества, которые этот процесс ускоряют. По типу действия ингибиторы можно разделить на обратимыеи необратимые. Удаление обратимых ингибиторов из системы (диализом, гельфильтрацией и др.) восстанавливает каталитическую активность фермента. Обратимо действуют эффекторы: 1. Близкие аналоги субстрата, которые связываются активным центром фермента, но не подвергаются превращению. Занимая активный центр, они препятствуют связыванию истинного субстрата, конкурируя с ним, и поэтому называются конкурентными ингибиторами. 2. Кофакторы ферментов, без которых апофермент вообще не обладает активностью. 3. Вещества, которые взаимодействуют с дополнительными, регуляторными центрами, несовпадающими с активным центром. Тем не менее, это взаимодействие изменяет конформацию в районе активного центра и влияет на кинетику ферментативного процесса. Такие соединения называются аллостерическими эффекторами. Они имеют важное биологическое значение, так как с их помощью осуществляется один из механизмов регуляции каталитической активности. Необратимую инактивацию вызывают соединения (найденные в живой природе или полученные путем синтеза), которые вступают в химическую реакцию с участком фермента, важным для проявления каталитической активности. Такие соединения, специфически реагирующие с определенными группами в молекулах ферментов (групп-специфические реагенты), используют для идентификации функциональных групп активного центра (метод химической модификации).
Активаторы ферментов - это вещества: 1) формирующие активный центр фермента ( Со2, Mg2, Zn2, Fe2, Ca2); 2) облегчающие образование фермент-субстратного комплекса ( Mg2); 3) восстанавливающие SH-группы ( глутатион, цистеин, меркаптоэтанол); 4) стабилизирующие нативную структуру белка-фермента.
Активаторами ферментов могут служить катионы и анионы разнообразных солей, способные к образованию различных комплексов как с самими ферментами, так и с их субстратами. Это возможно вследствие наличия в природе большого числа металлоферментов, содержащих тот или иной ион металла в активном центре ферментного белка. Механизмы активации энзиматических реакци, ; разнообразны
14. ТЕХНОЛОГИЯ ВЫДЕЛЕНИЯ И ОЧИСТКИ ФЕРМЕНТАТИВНЫХ ПРЕПОРАТОВ
Выделение и очистка фермента как из культуры микроорганизма (выращенного любым способом), так и из других природных источников весьма трудоемкая и дорогостоящая процедура, поэтому, если фермент можно использовать в виде неочищенного препарата, его не очищают. В промышленности широко применяют коммерческие препараты ферментов, чистота которых составляет всего 0,1 % (т.е. 99,9 % составляют примеси. Исходным материалом для получения препаратов ферментов служат: биомасса продуцента, фильтрат культуральной жидкости, экстракт из культуры микроорганизма или из тканей и органов растений и животных, из которых готовят препараты различной степени очистки.
Неочищенные ферментные препараты получают путем высушивания в мягком режиме культуры микроорганизмов вместе с остатками питательной среды. Такие препараты получают и путем упаривания экстракта из культуры продуцента, выращенного поверхностным способом, или из фильтрата культуральной жидкости (в случае глубинного выращивания микроорганизмов). Распространен также метод ацетоновых порошков, состоящий в осаждении и быстром обезвоживании при температуре не выше -10 °С тканей или вытяжек из них, содержащих ферменты. Технические препараты ферментов представляют собой либо высушенные до порошкообразного состояния продукты, либо жидкие концентраты, обычно характеризующиеся 50 %-м содержанием сухой массы веществ.
Для успешного выделения ферментов из клеточного содержимого необходимо очень тонкое измельчение исходного материала вплоть до разрушения субклеточных структур: лизосом, митохондрий, ядер и др. Для этого используют специальные мельницы и гомогенизаторы, а также ультразвук, метод попеременного замораживания и оттаивания ткани. Для высвобождения ферментов из мембранных структур клетки к гомогенатам добавляют небольшие количества детергентов или обрабатывают их энзимами — лизоцимом, целлюлазой, лецитиназой С. Особое внимание при выделении ферментов уделяют проведению всех операций в условиях, исключающих денатурацию белка (нейтральные значения рН, стабилизирующие добавки в виде белков, солей и специальных соединений).
На заключительных этапах очистки часто используют аффинную хроматографию (биоспецифическая хроматография, хроматография по сродству), которая основана на способности ферментов избирательно связывать те или иные лиганды — субстраты,
Очищенные ферментные препараты хранят при низкой температуре (до -80 °С). Для стабилизации ферментов в их препараты добавляют коферменты и субстраты.
Ферментационные технологии
При фермент. тех.можно использовать цельные живые клетки (микробов, клетки животных и растений) или клеточные компоненты (например, ферменты) с целью физ. Или хим. преобразований орг. вещ. Преимущества производства орг. продуктов биотех. способами перед чисто хим. методами достаточно многогранны:
• многие сложные орг. молекулы, такие, как белки и антибиотики, не могут практически быть синтезированы хим. способами;
• биоконверсия обеспечивает значительно больший выход целевого продукта;
• биологические системы функционируют при более низких температурах, менее высоких значениях рН (близких к нейтральному) и т. п.;
Сульфгидрилъные группы.Носитель хорошо удерживает фермент, если у обоих веществ есть сульфгидрильные группы. Они легко окисляются и образуют дисульфидные связи. Для этого достаточно подействовать на них кислородом воздуха
Включение в двухфазную средуОсобый случай иммобилизации – включение фермента в двухфазную среду. При этом фермент находится только в одной фазе, в которой он растворим. Как правило, такой фазой является водный раствор. Во второй фазе – органической – накапливается продукт реакции. Субстраты и продукты распределяются между обеими фазами. Меняя их соотношение, можно сдвигать равновесие в нужную сторону и тем самым управлять реакцией. Чтобы улучшить взаимодействие между фазами, одну из них диспергируют в другой, увеличивая поверхность раздела. После завершения реакции органическую фазу отделяют и извлекают из нее продукт. Оставшуюся фазу с ферментом можно использовать повторно. Двухфазные системы необходимы при работе с крупными молекулами субстратов, которые не могут взаимодействовать с мелкопористыми носителями.
18. ИММЩБИЛИЗАЦИЯ КЛЕТОК.
В 70-х годах XX века появились первые публикации об иммобилизации клеток микроорганизмов, а первое промышленное применение иммобилизованных клеток было осуществлено в Японии в 1974 г. С их помощью получали аспарагиновую кислоту.
Иммобилизованные клетки имеют ряд преимуществ как перед иммобилизованными ферментами, так и перед свободными клетками:
отсутствие затрат на выделение и очистку ферментов;
-снижение затрат на выделение и очистку продуктов реакции;
-более высокая активность и стабильность;
-возможность создания непрерывных и полунепрерывных автоматизированных процессов;
-способность к длительному функционированию полиферментных систем без экзогенных кофакторов.
Для иммобилизации могут быть использованы клетки в различном состоянии: живые и поврежденные в различной степени. Одностадийные реакции могут осуществлять и живые, и поврежденные клетки. Полиферментные реакции проводят с применением живых клеток, которые могут длительное время регенерировать АТФ и коферменты (НАДФ, НАД).
Проблема использования ферментативной активности иммобилизованных микроорганизмов имеет глубокие корни. Более 150 лет назад быстрый способ получения уксуса был основан на применении микроорганизмов, адсорбированных на древесной стружке. Методы иммобилизации клеток схожи с методами иммобилизации ферментов.
Химический метод основан на образовании ковалентных связей с активированным носителем, на поперечной сшивке клеток за счет активных групп в клеточной оболочке с бифункциональными реагентами (например, глутаровым альдегидом)
К физическим методам относятся адсорбция и агрегация.
Иммобилизация клеток путем включения в различные гели, мембраны, волокна основана на химических и физических взаимодействиях. Химические методы используются реже по сравнению с другими методами и малопригодны для иммобилизации живых клеток. Гораздо большее распространение получило включение клеток в состав гелей, мембран и волокон. При таком способе иммобилизации клетки могут сохранять жизнеспособность и в присутствии
21.АМИЛАЗЫ В ТЕХНОЛОГИИ ПИВОВАРЕНИЯ.
Задача пивоварения — получить высокий выход экстрактивных в-в из перерабатываемого сырья. Основным сырьём в пивоварении является ячменный солод. Выход экстракта находится в тесной взаимосвязи с активностью амилаз, образованных при солодораще-нии. Поскольку солодоращение с экономической точки зрения имеет ряд серьёзных недостатков, важной проблемой пивоваров является частичная замена солода несоложёным сырьём. С этой целью в затор вносят некоторое количество определённых ферментных препаратов, в т.ч. с амилазной активностью. Кроме того, ферменты в пивоварении применяются для стабилизации качества пива, улучшения его вкуса, аромата, стойкости при хранении.
В технологии спирта при переработке зерна хлебных злаков и картофеля основная задача — полностью перевести крахмал в сбраживаемые сахара. Для этого на спиртозаводах применяют бактериальную а-амилазу, а также глюкоамилазу. Последний фермент относительно кислото- и термостабилен и способен гид-ролизовать а-1,6-гликозидные связи. Поскольку гидролиз декстринов в сбраживаемый сахар является лимитирующей реакцией процесса брожения, то высокая активность глюкоамилазы обусловливает существенное сокращение длительности брожения. При 72-х часовом брожении расход а-амилазы должен составлять 1,5-2,0 ед АС, глюкоамилазы — 6,0-6,2 ед ГлА на 1 г перерабатываемого крахмала сырья. Препарат а-амилазы рекомендуется подавать в две точкитехнологической схемы: 0,5 ед АС на разжижение подвариваемой массы и 1 ед АС на 1 г крахмала при осахаривании вместе с глюкоамилазой.
|
6 МОЛЕКУЛЯРНЫЕ ЭФЕКТЫ ФЕРМЕНТАТИВНОГО КАТАЛИЗА
Механизмы ферментативного катализа определяются ролью функциональных групп активного центра фермента в химической реакции превращения субстрата в продукт. Выделяют 2 основных механизма ферментативного катализа: кислотно-основной катализ и ковалентный катализ. Кислотно-основной катализ Концепция кислотно-основного катализа объясняет ферментативную активность участием в химической реакции кислотных групп (доноры протонов) и/или основных групп (акцепторы протонов). Кислотно-основной катализ – часто встречающееся явление. Аминокислотные остатки, входящие в состав активного центра, имеют функциональные группы, проявляющие свойства как кислот, так и оснований. К аминокислотам, участвующим в кислотно-основном катализе, в первую очередь относят Цис, Тир, Сер, Лиз, Глу, Асп и Гис. Радикалы этих аминокислот в протонированной форме – кислоты (доноры протона), в депротонированной – основания (акцепторы протона). Благодаря этому свойству функциональных групп активного центра ферменты становятся уникальными биологическими катализаторами, в отличие от небиологических катализаторов, способных проявлять либо кислотные, либо основные свойства. Ковалентный катализ Ковалентный катализ основан на атаке нуклеофильных (отрицательно заряженных) или электрофильных (положительно заряженных) групп активного центра фермента молекулами субстрата с формированием ковалентной связи между субстратом и коферментом или функциональной группой аминокислотного остатка (как правило, одной) активного центра фермента. Действие сериновых протеаз, таких как трипсин, химотрипсин и тромбин, - пример механизма ковалентного катализа, когда ковалентная связь образуется между субстратом и аминокислотным остатком серина активного центра фермента. Термин «сериновые протеазы» связан с тем, что аминокислотный остаток серина входит в состав активного центра всех этих ферментов и участвует непосредственно в катализе. Рассмотрим механизм ковалентного катализа на примере химотрипсина, осуществляющего гидролиз пептидных связей при переваривании белков в двенадцатиперстной кишке. Субстратами химотрипсина служат пептиды, содержащие аминокислоты с ароматическими и циклическими гидрофобными радикалами (Фен, Тир, Три), что указывает на участие гидрофобныхp сил в формировании фермент-субстратного комплекса.
11 СЕЛЕКТИВНОСТЬ ФЕРМЕНТОВ
основаны на использовании хим. р-ций с участием ферментов. О содержании определяемого компонента судят либо по кол-ву конечного продукта ферментативной р-ции, либо, чаще, по начальной скорости процесса, положенного в основу методики определения Для наблюдения за скоростью ферментативной р-ции применяют обычно инструментальные методы, чаще других - люминесцентные, спектрофотометрич., электрохимические. Достоинства ферментативных методов анализа: высокая чувствительность, обусловленная активностью ферментов, природой индикаторных р-ций (с помощью к-рых определяют в-во) и способами детекции аналит. сигнала; высокая селективность и мягкие условия проведения анализа.
Определяемым компонентом в ферментативных методах анализа могут быть субстраты (в-ва, превращение к-рых катализирует фермент), сами ферменты, коферменты (в-ва, необходимые для осуществления каталитич. действияфермента) и эффекторы (соед., изменяющие каталитич. активность фермента - активаторы, ингибиторы).
Предел обнаружения, нижняя и верхняя границы определяемых содержаний компонентов зависят от кинетич. характеристик используемой индикаторной ферментативной р-ции и, прежде всего, каталитич. активности фермента.
Как правило, чувствительность определения ферментов, коферментов и эффекторов выше, чем чувствительность определения субстратов.
Высокая селективность ферментативных методов анализа обусловлена образованием фермент-субстратного комплекса в процессе каталитич. акта, требующим структурного соответствия активного центра фермента исубстрата. Поэтому большинство ферментов активно только в р-циях с субстратом одного определенного типа или с группой субстратов, имеющих общие структурные группы. Напр., фермент глюкозооксидаза катализирует окислениепрактически только одного вида глюкозы - 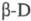 -глюкозу, к-рую можно определять без разделения сложной смеси моно-и олигосахаридов. В данном случае проявляется субстратная специфичность фермента.
Групповую специфичность можно наблюдать в случае действия альдегидоксидазы в р-циях превращения алифатич.альдегидов. Значительно менее селективны методы определения эффекторов, т. к. обычно имеется группа разл. соед., в той или иной степени меняющих каталитич. активность данного фермента. Однако селективностьопределения эффекторов м. б. и очень высокой.
Использование иммобилизованных ферментов.Недостатки ферментативных методов анализа обусловлены рядом особенностей ферментов: потерей функциональной активности и стабильности ферментов под воздействием разл. факторов; высокой стоимостью из-за невозможности многократного использования растворимыхферментов и трудности их выделения и очистки. Применение иммобилизованных ферментов расширило возможности ферментативных методов анализа. Более высокая стабильность и возможность многократного использования иммобилизованных ферментов позволили снизить стоимость анализов, повысить экспресс-ность, проводить хим. анализ в потоке
• каталитические биологические реакции намного специфичнее, чем реакции химического катализа;
• биологические процессы обеспечивают почти исключительно продукцию чистых изомеров одного типа, а не их смесей, как это часто бывает в реакциях хим. синтеза. Но вместе с тем био. способы в сравнении с хим. методами обладают рядом явных недостатков:
1. Био. системы могут легко быть загрязнены посторонней нежелательной микрофлорой.
2. Целевой продукт, синтезируемый био. способом, присутствует в довольно сложной смеси, что обусловливает необходимость разделения его от примеси ненужных веществ. 3. Биотех. производства требуют больших кол-в воды, которую в итоге необходимо удалять, сбрасывая вокружающую среду. 4. Биопроцессы обычно идут медленнее в сравнении со стандартными хим. Для каждого биотех. процесса должна быть разработана подходящая схема, а сам процесс должен постоянно наблюдаться и тщательно контролироваться. Для большинства практ. Биотех. процессов такими системами являются ферменторы или биореакторы, которые обеспечивают необходимые физ.условия, способствующие лучшему вз-вию катализатора со средой и поставляемым материалом. Биореакторы варьируют от простых сосудов до весьма сложных систем с различным уровнем компьютерного оснащения. Биореакторы изготавливаются в 2 вариантах (типах):
1) для нестерильных систем, когда нет абсолютной необходимости оперировать с чистыми кул-рами микроорг. (например, ферментация при пивоварении, производство пекарских дрожжей и т. п.).
2) предназначены для асептических процессов, обычно используемых в производстве таких соединений как, антибиотики, АК, полисахариды иодноклеточный бактериальный белок. Основное требование к биореакторам любого типа сводится кобеспечению оптим. условий роста продуцента или накоплению синтезируемого им продукта. Для достижения указанных целей нужно разрабатывать технологию, призванную оптимизировать процесс, а именно: использовать подходящий источник энергии, набор питательных веществ должен соответствовать пит. потребностям орг.-продуцента, из ростовой среды должны быть удалены соединения, ингибирующие егожизнедеятельность, должна быть подобрана соответствующая посевная доза и, наконец, обеспечены всеостальные требуемые физ-хим. условия. Эконом. рентабельные процессы в своей основе весьма сходны, независимо отизбранного продуцента, используемой среды и образуемого продукта. Главная задача – получение max кол-ва клеток содинаковыми св-вами при их выращивании в определенных тщательно контролируемых условиях. Фактически один и тот же биореактор может быть использован для производства ферментов, антибиотиков, орг.кислот или одноклеточного белка.
питательной среды размножаться в приповерхностных слоях гелей. Биокаталитическая активность целых иммобилизованных клеток в настоящее время может быть использована в различных отраслях науки и техники:
-при биосинтезе и трансформации таких соединений, как аминокислоты, органические кислоты, антибиотики, стероиды углеводы, углеводороды, нуклеотиды и нуклеозиды;
-в пивоварении и виноделии;
-при очистке сточных и природных вод;
-при извлечении металлов из сточных вод;
-при ассимиляции солнечной энергии;
-при изготовлении водородных солнечных элементов; -в азотфиксации;
-в аналитических целях при изготовлении электродов.
Наибольшее количество исследований по иммобилизации клеток микроорганизмов проведено японскими исследователями. Особые успехи были достигнуты ими в области синтеза аминокислот, органических кислот и антибиотиков. В Московском государственном университете был разработан метод получения аспарагиновой кислоты, который по эфективности не уступает японским. Клетки E.coli, включенные в армированный полиакриламидный гель, были с успехом использованы для получения аспарагиновой кислоты, период полужизни катализатора - 110 суток. Иммобилизовать можно не только клетки микроорганизмов, но и клетки растительных и животных тканей, используя их для синтеза физиологически активных соединений.
19. Получение глюкозно-фруктозных сиропов
Фруктоза самый сладкий сахар. Поэтому чем больше содержится фруктозы, тем слаще продукт при том же содержании сахара.
Для получения глюкозно-фруктозного сиропа в качестве исходного сырья используют в основном кукурузный крахмал. Содержание примесей в нем должно быть минимально, а содержание белка не должно превышать 0,4 %.
Для получения глюкозно-фруктозного сиропа используют гидролизаты крахмала с высоким содержанием глюкозы (96 %), полученные при ферментативном гидролизе крахмала. Для удаления растворимых примесей (зольные элементы, особенно ионы кальция, красящие вещества, протеин и др.) глюкозный сироп обрабатывают ионообменными смолами и активированным углем. Очищенный глюкозный сироп направляют на выпаривание до содержания СВ 40…50 %. При более высокой концентрации сиропа увеличивается его вязкость и падает скорость изомеризации. Иногда вместо выпаривания сироп стерилизуют при 125 ºС 2 мин, после чего его охлаждают до 60 ºС. В подготовленный субстрат добавляют ионы магния и кобальта для повышения активности фермента и бисульфит для предупреждения развития микрофлоры. Глюкоизомераза дозируется по его глюкоизомеразной активности. Процесс изомеризации длится около 20…24 ч до содержания фруктозы в гидролизате 42 %. Далее сироп отстаивают в течение нескольких часов и сливают так, чтобы осевший на дно фермент был покрыт слоем сиропа во избежание его контакта с воздухом. В реактор вновь подают свежий субстрат, и начинается новый цикл. Фермент используют от 5 до 30 дней и выводят из производства.
Полученный сироп подкисляют соляной кислотой до рН 4,5, очищают ионообменными -глюкозу, к-рую можно определять без разделения сложной смеси моно-и олигосахаридов. В данном случае проявляется субстратная специфичность фермента.
Групповую специфичность можно наблюдать в случае действия альдегидоксидазы в р-циях превращения алифатич.альдегидов. Значительно менее селективны методы определения эффекторов, т. к. обычно имеется группа разл. соед., в той или иной степени меняющих каталитич. активность данного фермента. Однако селективностьопределения эффекторов м. б. и очень высокой.
Использование иммобилизованных ферментов.Недостатки ферментативных методов анализа обусловлены рядом особенностей ферментов: потерей функциональной активности и стабильности ферментов под воздействием разл. факторов; высокой стоимостью из-за невозможности многократного использования растворимыхферментов и трудности их выделения и очистки. Применение иммобилизованных ферментов расширило возможности ферментативных методов анализа. Более высокая стабильность и возможность многократного использования иммобилизованных ферментов позволили снизить стоимость анализов, повысить экспресс-ность, проводить хим. анализ в потоке
• каталитические биологические реакции намного специфичнее, чем реакции химического катализа;
• биологические процессы обеспечивают почти исключительно продукцию чистых изомеров одного типа, а не их смесей, как это часто бывает в реакциях хим. синтеза. Но вместе с тем био. способы в сравнении с хим. методами обладают рядом явных недостатков:
1. Био. системы могут легко быть загрязнены посторонней нежелательной микрофлорой.
2. Целевой продукт, синтезируемый био. способом, присутствует в довольно сложной смеси, что обусловливает необходимость разделения его от примеси ненужных веществ. 3. Биотех. производства требуют больших кол-в воды, которую в итоге необходимо удалять, сбрасывая вокружающую среду. 4. Биопроцессы обычно идут медленнее в сравнении со стандартными хим. Для каждого биотех. процесса должна быть разработана подходящая схема, а сам процесс должен постоянно наблюдаться и тщательно контролироваться. Для большинства практ. Биотех. процессов такими системами являются ферменторы или биореакторы, которые обеспечивают необходимые физ.условия, способствующие лучшему вз-вию катализатора со средой и поставляемым материалом. Биореакторы варьируют от простых сосудов до весьма сложных систем с различным уровнем компьютерного оснащения. Биореакторы изготавливаются в 2 вариантах (типах):
1) для нестерильных систем, когда нет абсолютной необходимости оперировать с чистыми кул-рами микроорг. (например, ферментация при пивоварении, производство пекарских дрожжей и т. п.).
2) предназначены для асептических процессов, обычно используемых в производстве таких соединений как, антибиотики, АК, полисахариды иодноклеточный бактериальный белок. Основное требование к биореакторам любого типа сводится кобеспечению оптим. условий роста продуцента или накоплению синтезируемого им продукта. Для достижения указанных целей нужно разрабатывать технологию, призванную оптимизировать процесс, а именно: использовать подходящий источник энергии, набор питательных веществ должен соответствовать пит. потребностям орг.-продуцента, из ростовой среды должны быть удалены соединения, ингибирующие егожизнедеятельность, должна быть подобрана соответствующая посевная доза и, наконец, обеспечены всеостальные требуемые физ-хим. условия. Эконом. рентабельные процессы в своей основе весьма сходны, независимо отизбранного продуцента, используемой среды и образуемого продукта. Главная задача – получение max кол-ва клеток содинаковыми св-вами при их выращивании в определенных тщательно контролируемых условиях. Фактически один и тот же биореактор может быть использован для производства ферментов, антибиотиков, орг.кислот или одноклеточного белка.
питательной среды размножаться в приповерхностных слоях гелей. Биокаталитическая активность целых иммобилизованных клеток в настоящее время может быть использована в различных отраслях науки и техники:
-при биосинтезе и трансформации таких соединений, как аминокислоты, органические кислоты, антибиотики, стероиды углеводы, углеводороды, нуклеотиды и нуклеозиды;
-в пивоварении и виноделии;
-при очистке сточных и природных вод;
-при извлечении металлов из сточных вод;
-при ассимиляции солнечной энергии;
-при изготовлении водородных солнечных элементов; -в азотфиксации;
-в аналитических целях при изготовлении электродов.
Наибольшее количество исследований по иммобилизации клеток микроорганизмов проведено японскими исследователями. Особые успехи были достигнуты ими в области синтеза аминокислот, органических кислот и антибиотиков. В Московском государственном университете был разработан метод получения аспарагиновой кислоты, который по эфективности не уступает японским. Клетки E.coli, включенные в армированный полиакриламидный гель, были с успехом использованы для получения аспарагиновой кислоты, период полужизни катализатора - 110 суток. Иммобилизовать можно не только клетки микроорганизмов, но и клетки растительных и животных тканей, используя их для синтеза физиологически активных соединений.
19. Получение глюкозно-фруктозных сиропов
Фруктоза самый сладкий сахар. Поэтому чем больше содержится фруктозы, тем слаще продукт при том же содержании сахара.
Для получения глюкозно-фруктозного сиропа в качестве исходного сырья используют в основном кукурузный крахмал. Содержание примесей в нем должно быть минимально, а содержание белка не должно превышать 0,4 %.
Для получения глюкозно-фруктозного сиропа используют гидролизаты крахмала с высоким содержанием глюкозы (96 %), полученные при ферментативном гидролизе крахмала. Для удаления растворимых примесей (зольные элементы, особенно ионы кальция, красящие вещества, протеин и др.) глюкозный сироп обрабатывают ионообменными смолами и активированным углем. Очищенный глюкозный сироп направляют на выпаривание до содержания СВ 40…50 %. При более высокой концентрации сиропа увеличивается его вязкость и падает скорость изомеризации. Иногда вместо выпаривания сироп стерилизуют при 125 ºС 2 мин, после чего его охлаждают до 60 ºС. В подготовленный субстрат добавляют ионы магния и кобальта для повышения активности фермента и бисульфит для предупреждения развития микрофлоры. Глюкоизомераза дозируется по его глюкоизомеразной активности. Процесс изомеризации длится около 20…24 ч до содержания фруктозы в гидролизате 42 %. Далее сироп отстаивают в течение нескольких часов и сливают так, чтобы осевший на дно фермент был покрыт слоем сиропа во избежание его контакта с воздухом. В реактор вновь подают свежий субстрат, и начинается новый цикл. Фермент используют от 5 до 30 дней и выводят из производства.
Полученный сироп подкисляют соляной кислотой до рН 4,5, очищают ионообменными
|