От вражды к дружбе
· Транспозоны (как и вирусы) изначально, конечно, возникли как геномные паразиты.
· Закон жизни: как только где-то возникает что-то похожее на кооперацию, сотрудничество, взаимопомощь – в такой системе немедленно заводятся паразиты.
· Геном – это содружество генов, помогающих друг другу размножаться. В нем не могли не завестись паразиты – гены, пользующиеся услугами других генов для своего размножения, но ничего не дающие взамен.
· Под действием отбора отношения паразита с хозяйским геномом может пойти по одному из двух путей: антагонистическому («гонка вооружений») или кооперативному («молекулярное одомашнивание»). Широко распространено и то, и другое.
· Для «одомашнивания» нужно, чтобы в генетическом паразите произошла случайная мутация, из-за которой он (или какой-нибудь его фрагмент) стал бы чем-то полезен хозяйскому геному.
· Два ярких примера одомашнивания: 1) Белки, осуществляющие VDJ-рекомбинацию (они называются RAG-белки) – это «одомашненные» транспозазы, чьи гены некогда были частью ДНК-овых транспозонов. 2) Белки, осуществляющие геномную перестройку в МАК-геноме инфузорий – это тоже прирученные транспозазы.
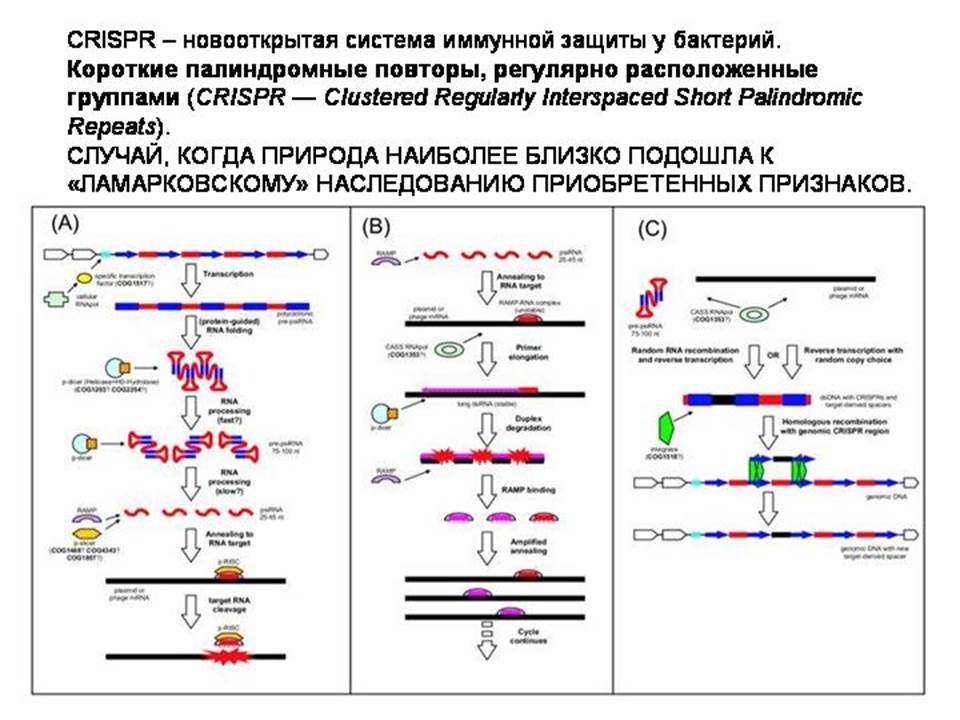
К вопросу о гонке вооружений между паразитами и хозяевами.
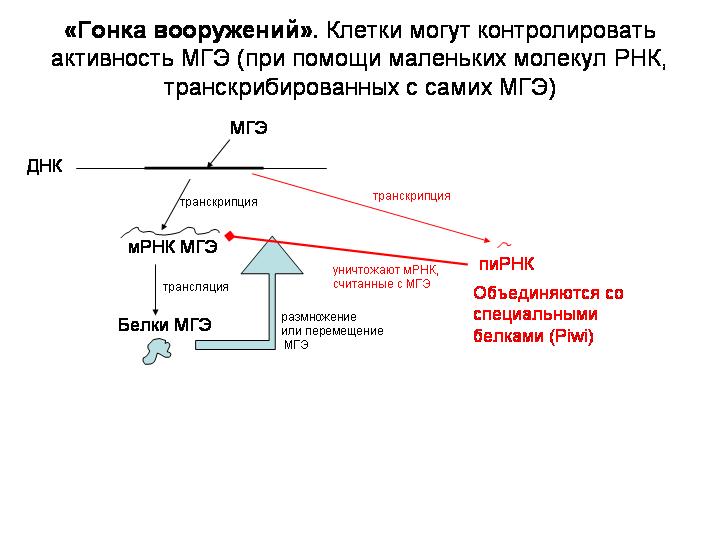
Живым организмам не выгодно, чтобы транспозоны постоянно прыгали в их геноме, вызвая многочисленные (и в основном вредные) мутации. Поэтому отбор должен поддерживать у организмов такие мутации, которые помогут обуздывать активность МГЭ. И действительно, недавно была открыта специальная молекулярная система, позволяющие клетке предотвращать прыжки МГЭ.
Центральную роль в этой системе играют маленькие молекулы РНК, считанные с самих МГЭ (пиРНК, длиной 24-29 нуклеотидов). Эти пиРНК объединяются в комплексы со специальными белками (Piwi). Эти комплексы затем находят молекулы мРНК, считанные с МГЭ, и уничтожают их, тем самым не давая МГЭ прыгать и размножаться.
Они режут мРНК МГЭ на кусочки, причем некоторые из этих кусочков сами потом могут служить в качестве пиРНК. Поэтому получается нечто вроде цепной реакции.
Очень важный момент: поскольку есть специальные системы подавления активности МГЭ, это значит, что в каких-то неблагоприятных, стрессовых условиях эти системы могут давать сбои, ломаться. И тогда МГЭ должны начинать прыгать бесконтрольно. Так оно, по-видимому, и есть. Показано, что МГЭ могут активизироваться в стрессовых условиях. И получается, нечто подобное системе повышения скорости мутирования при стрессе, которая есть у бактерий, и о которой мы уже говорили. Когда жизнь становится очень плохой, системы контроля МГЭ начинают давать сбои, и это приводит к росту мутагенеза, вызванного активностью МГЭ.
Примеры одомашнивания:
· RAG-белки (осуществляют «нарезку» ДНК в ходе VDJ-рекомбинации) – произошли от транспозаз
· Теломеразы (восстанавливают кончики хромосом, укорачивающиеся после каждого деления) – произошли от обратных транскриптаз
· Фрагменты МГЭ часто становятся регуляторными элементами хозяйского генома. Значительная часть (как минимум 16%) регуляторных элементов (в т.ч. промоторов, энхансеров) в геномах плацентарных млекопитающих представляют собой бывшие фрагменты МГЭ
· Ряд белков вирусного происхождения работает в плаценте (функции: 1) слияние клеток, 2) защита эмбриона от иммунной системы матери, 3) защита эмбриона от вирусов).
· Многие МГЭ «по совместительству» служат генами маленьких регуляторных РНК. Эти РНК используются клеткой прежде всего для того, чтобы контролировать активность самих МГЭ, но не только. Они могут выполнять и другие регуляторные функции: например, своевременно уничтожать материнские мРНК в развивающемся эмбрионе

Рассмотрим один яркий пример «молекулярного одомашнивания» из жизни насекомых.
Наездники — одна из самых разнообразных групп насекомых, включающая десятки тысяч видов. Они играют огромную роль в природе, а также в сельском хозяйстве, контролируя численность растительноядных насекомых, в том числе многих вредителей. Личинки наездников развиваются в теле жертвы, пожирая ее заживо. В свое время эта необычайная «жестокость природы» повлияла на мировоззрение Чарльза Дарвина, который заметил в одном из писем, что не может себе представить, как мог благой и милосердный Создатель сотворить наездников, чтобы они заживо пожирали своих жертв (причем сотворить не единожды, а в десятках тысяч вариаций, как мы могли бы сейчас добавить).
Многие наездники впрыскивают в жертву, кроме своих яиц, также и особые вирусоподобные частицы. Оболочка этих вирусоподобных частиц очень похожа на оболочку других вирусов насекомых, однако генетическая «начинка» у них совсем другая. Эти частицы называют «поли-ДНК-вирусами», или ПДВ.

Каждый ПДВ содержит несколько маленьких кольцевых молекул ДНК с генами белков, подавляющих защитную (иммунную) реакцию организма хозяина (например, гусеницы). Это позволяет личинкам наездника беспрепятственно развиваться в теле жертвы.
При этом, в отличие от всех нормальных вирусов, ПДВ не содержат специфических вирусных генов, необходимых для размножения. Поэтому ПДВ не могут размножаться в организме гусеницы. Не размножаются они и в организме наездника. По сути дела, они вообще сами не размножаются. Новые ПДВ образуются только в яичниках самки наездника. Яичники синтезируют ПДВ точно так же, как любой орган многоклеточного животного синтезирует различные вещества и молекулярные комплексы для внутреннего использования или выведения наружу.
Эти удивительные особенности поли-ДНК-вирусов заставляли некоторых экспертов сомневаться в их вирусной природе. Может быть, ПДВ в действительности являются не вирусами, а просто некими молекулярными комплексами, которые возникли в ходе эволюции наездников как одна из адаптаций к паразитизму? Может быть, их внешнее сходство с вирусами случайно?
В 2009 году генетики из Франции и Швейцарии показали, что ПДВ все-таки происходят от настоящего вируса, который некогда встроился в геном предков наездников и подвергся «молекулярному одомашниванию».
«Геном» изученных ПДВ, то есть тот генетический материал, который находится в вирусоподобной частице, состоит из 30 кольцевых двухцепочечных ДНК. Эти молекулы ДНК кодируют белки-иммуносупрессоры, совершенно не похожие на белки известных вирусов. Эти кольцевые «хромосомы» ПДВ изготавливаются из копий фрагментов геномной ДНК самого наездника.
Гены, необходимые для образования самого ПДВ, в том числе гены белков оболочки этого псевдовируса, отсутствуют в его «геноме». Именно это и вызвало сомнения в том, что ПДВ – потомки диких вирусов. Нужно было найти гены, ответственные за построение вирусных частиц. И эти гены удалось найти: в геноме наездника нашли несколько встроенных вирусных генов, «работающих», не испорченных мутациями. Удалось доказать, что именно эти гены явно вирусного происхождения, встроенные в геном наездника, и отвечают за создание вирусных частич ПДВ в яичниках самки. Именно в них закодированы белки оболочки ПДВ.
Таким образом, когда-то в геном предков наездников встроился вирус, который затем подвергся «одомашниванию». Вирусные гены впоследствии рассеялись по геному наездника, перестали включаться в состав вирусной частицы, но по-прежнему продолжают работать, обеспечивая сборку этих частиц, которые наполняются теперь совсем другой генетической «начинкой».
Иначе говоря, встроившийся вирус «пошел на запчасти», его гены стали необходимой частью генома наездника.
Сравнив между собой гены множества разных наездников, авторы пришли к выводу, что около 100 млн лет назад (в середине мелового периода, когда происходила быстрая сопряженная диверсификация цветковых растений и насекомых) в геном какого-то древнего наездника встроился вирус, который вскоре был одомашнен и стал помогать наездникам справляться с иммунной системой гусениц. Симбиоз с вирусом оказался настолько выгодным, что потомки этого наездника необычайно размножились и дали начало 17 тысячам современных видов.
Уже на самых ранних этапах эволюции симбиотического комплекса генетическая «начинка» вирусных частиц была заменена: теперь вместо вирусных генов в них стали упаковываться гены наездника.
Это исследование еще раз показало, что симбиоз — в том числе симбиоз животных с вирусами — является одним из важнейших путей формирования эволюционных новшеств.
И это не конец истории!

Недавно был обнаружен еще один поразительный факт.
Многие насекомые буквально нашпигованы различными симбиотическими микробами. Бактерии-симбионты помогают своим хозяевам решать многие жизненные задачи: от синтеза незаменимых аминокислот и витаминов до переваривания химически чистой целлюлозы, фиксации атмосферного азота и борьбы с сорняками на грибных плантациях.
В круг задач, выполняемых бактериями — симбионтами насекомых, входит и защита хозяев от наездников. Именно эту роль взяла на себя бактерия Hamiltonella defensa, обитающая в клетках многих насекомых, в том числе тлей. Гамильтонелла, как и многие другие внутриклеточные симбионты, наследуется по материнской линии в ряду поколений насекомых-хозяев (бактерия проникает в яйца, откладываемые зараженной самкой).
Ранее было замечено, что разновидности гамильтонеллы различаются по своим защитным свойствам: одни обеспечивают надежную тли от наездника, убивая до 80–90% личинок паразита, тогда как другие защищают хозяина гораздо хуже или не защищают вовсе. Кроме того, обнаружилось, что защитные свойства гамильтонеллы коррелируют с зараженностью бактерии вирусом-бактериофагом APSE (A. pisum secondary endosymbiont). Известно несколько разновидностей этого вируса, причем в геноме каждой разновидности вируса содержатся гены белков, токсичных для животных. Было высказано предположение, что именно эти токсины, закодированные в вирусном геноме, используются симбиотическим комплексом «тля—бактерия» для борьбы с общим врагом — личинками наездника. Это предположение косвенно подтвердилось в ходе сравнения защитных свойств трех штаммов гамильтонеллы, два из которых имеют в своем геноме встроенный геном разновидности вируса APSE-3, а третий не заражен вирусом. Оказалось, что штаммы с вирусом обеспечивают очень высокий уровень защиты тли от наездника, а штамм без вируса не обладает этим свойством.
Этих данных, однако, было недостаточно для того, чтобы считать роль вируса APSE в защите тли от наездника доказанной. Дело в том, что в данном симбиотическом комплексе участвуют трое — тля, бактерия и вирус; у каждого из трех компонентов есть свой собственный геном, причем каждый из трех геномов существует в виде множества вариаций. Поэтому, например, если комплекс «тля + бактерия + вирус» защищен от паразитов, а комплекс «тля + бактерия» — нет, то причиной может быть не только присутствие или отсутствие вируса, но и генетические различия между штаммами (линиями) бактерий и тлей. В принципе, наличие вируса в геноме тех штаммов бактерии, которые защищают тлю от наездников, может оказаться случайным совпадением.
Чтобы строго доказать роль вируса во взаимовыгодном сотрудничестве бактерии и тли, нужно было получить генетически чистые линии тлей и бактерий, различающиеся только наличием или отсутствием в бактериальном геноме встроенного фага APSE.

Именно это и сделали в 2009 году американские энтомологи. Им удалось вывести три линии генетически идентичных тлей: 1) с гамильтонеллой, зараженной вирусом APSE, 2) с точно такой же гамильтонеллой, но без вируса; 3) без гамильтонеллы. Этих тлей отдавали «на растерзание» наездникам, которые откладывали в них свои яйца. Затем ученые подсчитывали в каждой из трех линий процент выживших личинок наездника (который равен проценту погибших тлей, так как в каждую тлю наездник откладывает одно яйцо, после чего выжить может только кто-то один: либо тля, либо личинка наездника).
В первой из трех линий личинки наездника сумели погубить лишь около 6% тлей, во второй и третьей — около 80%. Эти результаты доказывают, что вирус APSE действительно необходим для защиты тли от наездника.
Интересно, что содержание в своем теле защитных симбионтов не является «бесплатным удовольствием» ни для тлей, ни для бактерий. При выращивании в лаборатории в отсутствие наездников тли, не зараженные гамильтонеллой, размножаются успешнее зараженных, и в результате со временем популяция тлей может полностью освободиться от бактерии. То же самое происходит и в паре «бактерия—вирус»: оказалось, что гамильтонеллы в лабораторных линиях полностью освобождаются от вируса за 3–4 года. Разумеется, ситуация меняется на обратную, если тли регулярно подвергаются нападению наездников. В этом случае естественный отбор благоприятствует как распротранению гамильтонеллы в популяции тлей, так и распространению вируса APSE в популяции гамильтонелл.
Таким образом, в «эволюционной гонке вооружений» наездников и их жертв симбиотические вирусы участвуют в игре на обеих сторонах. Фактически, насекомые ведут друг с другом настоящую «вирусологическую войну».
Исследования подобных сибмиотических систем только начинаются. Мы еще очень мало знаем о симбиотических вирусах.
Не исключено, что та поразительная скорость, с которой насекомые приспосабливаются к новым условиям (например, к новым кормовым растениям) или вырабатывают устойчивость к ядам, отчасти объясняется деятельностью вирусов, проводящих разнообразные генно-инженерные эксперименты на бактериях — симбионтах насекомых.
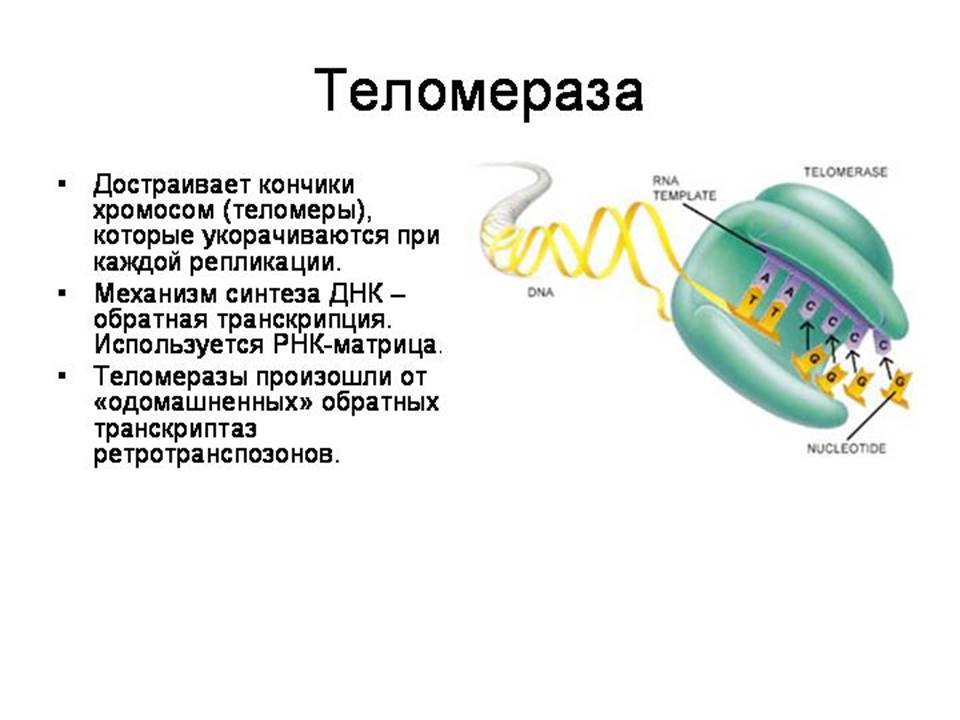
Укорочение линейных хромосом при репликации. Оловников и Нобелевская премия. Шаблонный участок TERC содержит последовательность 3'-ЦААУЦЦЦААУЦ-5'. Теломераза связывает первые несколько нуклеотидов шаблона с последней теломерной последовательностью на хромосоме, добавляет новый повторяющийся участок (5'-ГГТТАГ-3'), отделяется, связывает новый 3'-конец теломеры с шаблоном и повторяет весь процесс заново.