У РАСТЕНИЙ
«...Способность растения к оптимальной устойчивости отражает меру его надежности».
 Д. М. Гродзинский
Д. М. Гродзинский
Способность к защите от действия неблагоприятных абиотических и биотических факторов среды — столь же обязательное свойство любого организма, как питание, движение, размножение и др. Эта функция появилась одновременно с возникновением первых живых организмов и в ходе дальнейшей эволюции развивалась и совершенствовалась. Поскольку повреждающих и уничтожающих факторов множество, возникшие способы защиты от них оказались самыми разными — от метаболических механизмов до морфологических приспособлений (колючек и др.). Выживаемость и расселение по новым экологическим нишам определялись способностью организмов приспосабливаться к необычным условиям среды. Адаптация, т. е. приспособление организма к конкретным условиям существования, у индивидуума достигается за счет физиологических механизмов (физиологическая адаптация), а у популяции организмов (вида) — благодаря механизмам генетической изменчивости и наследственности (генетическая адаптация).
Если нарушения обмена веществ и функциональной активности определять как «отказ», то в физиологии растений можно использовать технический термин «надежность», подразумевая под этим безотказность функционирования растительного организма в нормальных условиях существования и при отклонении от нормы. Надежность растительного организма определяется его способностью не допускать или ликвидировать отказы на разных уровнях: молекулярном, субклеточном, клеточном, тканевом, органном, организменном и популяционном. Для предотвращения отказов используются системы стабилизации: принцип избыточности, принцип гетерогенности равнозначных компонентов, механизмы гомеостаза. Для ликвидации возникших отказов служат системы репарации (восстановления). На каждом уровне биологической организации действуют свои механизмы. На молекулярном уровне принцип избыточности находит свое выражение, например, в полиплоидии, на организменном — в образовании большого количества гамет и семян. Примерами восстановительной активности на молекулярном уровне служит энзиматическая репарация поврежденной ДНК, на организменном - пробуждение пазушных почек при повреждении апикальной Меристемы, регенерация и т. д.
 Защита от неблагоприятных факторов среды у растений обеспечивается особенностями анатомического строения (кутикула, корка, механические ткани и т. д.), специальными органами защиты (жгучие волоски, колючки), двигательными и физиологическими реакциями, выработкой защитных веществ (смол, фитоалексинов, фитонцидов, токсинов, защитных белков).
Защита от неблагоприятных факторов среды у растений обеспечивается особенностями анатомического строения (кутикула, корка, механические ткани и т. д.), специальными органами защиты (жгучие волоски, колючки), двигательными и физиологическими реакциями, выработкой защитных веществ (смол, фитоалексинов, фитонцидов, токсинов, защитных белков).
 Надежность организма проявляется в эффективности его защитных приспособлений, в его устойчивости к действию неблагоприятных факторов внешней среды: высокой и низкой температуры, недостатка кислорода, дефицита воды, засоления и загазованности среды, ионизирующих излучений, инфекции и др. Эти неблагоприятные факторы в последнее время часто называют стрессорами, а реакцию организма на любые отклонения от нормы — стрессом. Самые разнообразные неблагоприятные факторы могут действовать длительное время или оказывают сравнительно кратковременное, но сильное влияние. В первом случае, как правило, в большей степени проявляются специфические механизмы устойчивости, во втором — неспецифические. Учение о неспецифических ответах клеток на воздействие разнообразных факторов внешней среды было разработано Н. Е. Введенским, Д. Н. Насоновым и В. Я. Александровым. Проблема надежности в физиологии растений во всем объеме поставлена и развита Д. М. Гродзинским (1983).
Надежность организма проявляется в эффективности его защитных приспособлений, в его устойчивости к действию неблагоприятных факторов внешней среды: высокой и низкой температуры, недостатка кислорода, дефицита воды, засоления и загазованности среды, ионизирующих излучений, инфекции и др. Эти неблагоприятные факторы в последнее время часто называют стрессорами, а реакцию организма на любые отклонения от нормы — стрессом. Самые разнообразные неблагоприятные факторы могут действовать длительное время или оказывают сравнительно кратковременное, но сильное влияние. В первом случае, как правило, в большей степени проявляются специфические механизмы устойчивости, во втором — неспецифические. Учение о неспецифических ответах клеток на воздействие разнообразных факторов внешней среды было разработано Н. Е. Введенским, Д. Н. Насоновым и В. Я. Александровым. Проблема надежности в физиологии растений во всем объеме поставлена и развита Д. М. Гродзинским (1983).
Канадский ученый Г. Селье со второй половины 30-х годов ввел в медицину понятие «стресс» (от англ, stress — напряжение). По Селье, стресс — это совокупность всех неспецифических изменений, возникающих в организме животного под влиянием любых сильных воздействий (стрессоров), включающих перестройку защитных сил организма. Эта перестройка сопровождается увеличением в крови адреналина и других гормонов, мобилизующих обмен веществ. По Селье, стресс как реакция организма на неблагоприятное воздействие проходит три фазы: 1) тревоги, 2) резистентности (адаптации) и 3) истощения. Если при сильных воздействиях последняя фаза наступает и развивается быстро, то организм погибает.
Перенос теории стресса в том виде, как это изложено выше, на растительные объекты кажется на первый взгляд дискуссионным. У растений нет ни нервной системы, ни тех гормонов, которые участвуют в стрессовых реакциях у животных. Однако если рассматривать не частности, а суть теории стресса, как неспецифической реакции клетки и организма в целом на экстремальные воздействия, то этот вопрос в физиологии растений заслуживает самого пристального внимания, хотя и требует определенных корректив.
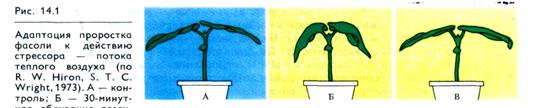
Во-первых, применительно к растениям 1-я фаза стрессовой реакции не может быть названа фазой тревоги. Для растений можно говорить о следующих трех фазах: 1) первичной стрессовой реакции, 2) адаптации, 3) истощения ресурсов надежности. На рис. 14.1 наглядно демонстрируются две первые фазы стресса у растений. Листья проростков фасоли, обдуваемые феном (38 °С), через 12 — 30 мин опускаются вследствие завядания (фаза первичной реакции), но затем снова поднимаются (фаза адаптации), несмотря на продолжающееся действие «суховея».
 Во-вторых, растительные организмы в отличие от животных в большинстве случаев реагируют на стрессор не активацией обмена веществ, а, наоборот, снижением своей функциональной активности. В связи с этим при стрессе в тканях растений возрастает концентрация гормонов, тормозящих обмен веществ, — этилена и АБК, о чем более подробно будет сказано ниже.
Во-вторых, растительные организмы в отличие от животных в большинстве случаев реагируют на стрессор не активацией обмена веществ, а, наоборот, снижением своей функциональной активности. В связи с этим при стрессе в тканях растений возрастает концентрация гормонов, тормозящих обмен веществ, — этилена и АБК, о чем более подробно будет сказано ниже.
Факторы, способные вызвать стресс у растительных организмов, можно подразделить на три основные группы:
а) физические: недостаточная или избыточная влажность, освещенность или температура, радиоактивное излучение, механические воздействия;
б) химические: соли, газы, ксенобиотики (гербициды, инсектициды, фунгициды, промышленные отходы и др.);
в) биологические (поражение возбудителями болезней или вредителями, конкуренция с другими растениями, влияние животных, цветение, созревание плодов).
Действие одного и того же фактора с одним и тем же уровнем интенсивности может вызывать или не вызывать стресс у растения в зависимости от его сопротивляемости. Так, по отношению к засухе растения делятся на две группы: 1) пойкило-гидрические, не регулирующие свой водный режим и допускающие большую потерю воды (до воздушно-сухого состояния), не теряя жизнеспособности; 2) гомойогидрические растения, регулирующие водный обмен и отвечающие стрессом на водный дефицит.
Устойчивость растения к стрессовому воздействию зависит и от фазы онтогенеза. Наиболее устойчивы растения, находящиеся в покоящемся состоянии (в виде семян, луковиц и т. п.). Наиболее чувствительны — растения в молодом возрасте, в период появления всходов, так как в условиях стресса прежде всего повреждаются те звенья метаболизма, которые связаны с активным ростом. Затем по мере роста и развития устойчивость растений к стрессовым воздействиям постепенно возрастает вплоть до созревания семян. Однако период формирования гамет также является критическим, поскольку растения в это время высокочувствительны к стрессу и реагируют на действие стрессоров снижением продуктивности.
1 От греч. «хепох» — чужой.
 Механизмы стресса на клеточном уровне.Впервые стереотипные реакции животных клеток на различные воздействия были изучены Н. Е. Введенским (учение о парабиозе) и Д. Н. Насоновым и В. Я. Александровым (учение о паранекрозе). Н. Е. Введенский (1901), использовав внеклеточные электроды, установил, что при действии слабых раздражителей клетка отвечает временной электропозитивацией, а при более сильном воздействии развивается злектронегативация, что соответствует деполяризации мембранного потенциала. Насонов и Александров (1940) показали, что при разнообразных слабых воздействиях в клетках снижаются сорбция красителей, свето-рассеивание цитоплазмы и ее вязкость. Наоборот, сильное раздражение вызывает в клетке увеличение сорбции красителей, повышение светорассеивания и вязкости цитоплазмы.
Механизмы стресса на клеточном уровне.Впервые стереотипные реакции животных клеток на различные воздействия были изучены Н. Е. Введенским (учение о парабиозе) и Д. Н. Насоновым и В. Я. Александровым (учение о паранекрозе). Н. Е. Введенский (1901), использовав внеклеточные электроды, установил, что при действии слабых раздражителей клетка отвечает временной электропозитивацией, а при более сильном воздействии развивается злектронегативация, что соответствует деполяризации мембранного потенциала. Насонов и Александров (1940) показали, что при разнообразных слабых воздействиях в клетках снижаются сорбция красителей, свето-рассеивание цитоплазмы и ее вязкость. Наоборот, сильное раздражение вызывает в клетке увеличение сорбции красителей, повышение светорассеивания и вязкости цитоплазмы.
К первичным неспецифическим процессам, происходящим в клетках растений при сильном и быстро нарастающем действии стрессора, относятся следующие:
1. Повышение проницаемости мембран, деполяризация мембранного потенциала плазмалеммы.
2. Вход Са2+ в цитоплазму (из клеточных стенок и внутриклеточных компартментов: вакуоли, ЭС, митохондрий).
3. Сдвиг рН цитоплазмы в кислую сторону.
4. Активация сборки актиновых микрофиламентов и сетей цитоскелета, в результате чего возрастает вязкость и свето-рассеивание цитоплазмы.
5. Усиление поглощения О2, ускоренная трата АТР, развитие свободнорадикальных реакций.
6. Возрастание гидролитических процессов.
7. Активация и синтез стрессовых белков (см. ниже).
8. Усиление активности Н+-помпы в плазмалемме (и, возможно, в тонопласте), препятствующей неблагоприятным сдвигам ионного гомеостаза.
9. Увеличение синтеза этилена и АБК, торможение деления и роста, поглотительной активности клеток и других физиологических и метаболических процессов, осуществляющихся в обычных условиях. Торможение функциональной активности клеток происходит в результате действия ингибиторов и переключения энергетических ресурсов на преодоление неблагоприятных сдвигов.
Перечисленные стрессовые реакции наблюдаются при действии любых стрессоров. Они направлены на защиту внутриклеточных структур и устранение неблагоприятных изменений в клетках. Все эти явления адаптационного синдрома (стресса) взаимосвязаны и развиваются как каскадные процессы. В настоящее время усилия направлены на полную расшифровку механизмов стресса на молекулярном и клеточном уровнях. Однако необходимо помнить, что наряду с неспецифическим эффектом все стрессоры оказывают и специфическое воздействие на клетки и ткани.
Особый интерес вызывают данные об активации в клетках в условиях стресса синтеза так называемых стрессовых белков с одновременным ослаблением синтеза белков, образующихся в нормальных условиях. Так, у многих растений выявлены белки теплового шока. Например, у кукурузы синтез этих белков индуцируется температурой 45 С. Гены температурного шока лишены нитронов, мРНК имеет полупериод жизни 2 ч, а белки — около 20 ч, в течение которого клетка сохраняет терморе-зистентность. Показано, что некоторые из этих белков предсу-ществуют в цитоплазме и в условиях стресса активируются фрагментированием. В ядре и в ядрышке белки теплового шока образуют гранулы, связывая матрицы хроматина, необходимые для нормального метаболизма. После прекращения стрессового состояния эти матрицы вновь освобождаются и начинают функционировать. Один из белков теплового шока стабилизирует плазмалемму, проницаемость которой для внутриклеточных веществ в условиях стресса возрастает.
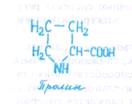 Кроме синтеза шоковых белков, показывающего, что в ге-номе записана специальная программа, связанная с переживанием стресса, при неблагоприятных обстоятельствах в клетках возрастает содержание углеводов, пролина, которые участвуют в защитных реакциях, стабилизируя цитоплазму. При водном дефиците и засолении у ряда растений (ячмень, шпинат, хлопчатник и др.) концентрация пролина в цитоплазме возрастает в 100 раз и более. Благодаря своим гидрофильным группам пролин может образовывать агрегаты, которые ведут себя как гидрофильные коллоиды. Этим объясняется высокая растворимость пролина, а также способность его связываться с поверхностными гидрофильными остатками белков. Необычный характер взаимодействия агрегатов пролина с белками повышает растворимость белков и защищает их от денатурации. Накопление пролина как осмотически активного органического вещества благоприятствует удержанию воды в клетке.
Кроме синтеза шоковых белков, показывающего, что в ге-номе записана специальная программа, связанная с переживанием стресса, при неблагоприятных обстоятельствах в клетках возрастает содержание углеводов, пролина, которые участвуют в защитных реакциях, стабилизируя цитоплазму. При водном дефиците и засолении у ряда растений (ячмень, шпинат, хлопчатник и др.) концентрация пролина в цитоплазме возрастает в 100 раз и более. Благодаря своим гидрофильным группам пролин может образовывать агрегаты, которые ведут себя как гидрофильные коллоиды. Этим объясняется высокая растворимость пролина, а также способность его связываться с поверхностными гидрофильными остатками белков. Необычный характер взаимодействия агрегатов пролина с белками повышает растворимость белков и защищает их от денатурации. Накопление пролина как осмотически активного органического вещества благоприятствует удержанию воды в клетке.
Неспецифический характер некоторых реакций клеток на стресс позволяет мобилизовать резервные возможности организма для общего быстрого ответа на действие неблагоприятных факторов окружающей среды, в том числе и при воздействии необычных раздражителей. В невысоких дозах повторяющиеся стрессы способствуют закаливанию организма, причем во многих случаях показано, что закаливание по отношению к одному стрессорному фактору способствует повышению устойчивости организма и к некоторым другим стрессорам.
Механизмы стресса и адаптации на организменном уровне.На разных уровнях организации приспособление к экстремальным условиям осуществляется у растений неодинаково. Чем выше уровень биологической организации (клетка, организм, популяция), тем большее число механизмов одновременно участвует в адаптации растений к стрессовым воздействиям.
 На организменном уровне сохраняются все механизмы адаптации, свойственные клетке, но дополняются новыми, отражающими взаимодействие органов в целом растении. Прежде всего это конкурентные отношения между органами за физиологически активные вещества и трофические факторы. Эти отношения построены на силе аттрагирующего (притягивающего) действия. Подобный механизм позволяет растениям в экстремальных условиях сформировать лишь такой минимум генеративных органов (аттрагирующих центров), которые они в состоянии обеспечить необходимыми веществами для нормального созревания. Например, при неблагоприятных условиях в колосе злака формируются не все семена, а лишь немногие, но эти оставшиеся достигают обычных размеров. Точно так же у плодовых деревьев в результате конкуренции за питательные вещества между ранее и позже заложившимися плодами часть из них опадает и тем в большей степени, чем хуже условия существования растения в целом. При неблагоприятных условиях резко ускоряются процессы старения и опадения нижних листьев, причем продукты их распада используются для питания более молодых органов.
На организменном уровне сохраняются все механизмы адаптации, свойственные клетке, но дополняются новыми, отражающими взаимодействие органов в целом растении. Прежде всего это конкурентные отношения между органами за физиологически активные вещества и трофические факторы. Эти отношения построены на силе аттрагирующего (притягивающего) действия. Подобный механизм позволяет растениям в экстремальных условиях сформировать лишь такой минимум генеративных органов (аттрагирующих центров), которые они в состоянии обеспечить необходимыми веществами для нормального созревания. Например, при неблагоприятных условиях в колосе злака формируются не все семена, а лишь немногие, но эти оставшиеся достигают обычных размеров. Точно так же у плодовых деревьев в результате конкуренции за питательные вещества между ранее и позже заложившимися плодами часть из них опадает и тем в большей степени, чем хуже условия существования растения в целом. При неблагоприятных условиях резко ускоряются процессы старения и опадения нижних листьев, причем продукты их распада используются для питания более молодых органов.
Важнейший и очень характерный для растений механизм защиты от последствий действия экстремальных факторов — процесс замены поврежденных или утраченных органов путем регенерации и роста пазушных почек. Во всех этих процессах коррелятивного роста участвуют межклеточные системы регуляции (гормональная, трофическая и электрофизиологическая).
При неблагоприятных условиях существования в растениях резко возрастает выработка этилена и АБК, снижающих обмен веществ, тормозящих ростовые процессы, способствующих старению и опадению органов, переходу растительного организма в состояние покоя. Одновременно в тканях снижается содержание ауксина, цитокинина и гиббереллинов. Эта стереотипная реакция гормональной системы на экстремальные условия очень характерна для растительных организмов. Здесь наблюдается явное соответствие теории стресса, предложенной Селье для животных, с той только разницей, что у растений в условиях стресса ведущую роль играют фитогормоны, тормозящие их функциональную активность.
Стресс на популяционном уровне. Вусловиях длительного и сильного стресса в период истощения гибнут те индивидуумы, у которых генетически норма реакции на данный эк стремальный фактор ограничена узкими пределами. Эти растения устраняются из популяции, а семенное потомство образуют лишь генетически более устойчивые растения. В результате общий уровень устойчивости в популяции возрастает. Таким образом, на популяционном уровне в стрессовую реакцию включается дополнительный фактор — отбор, приводящий к появлению более приспособленных организмов и новых видов (генетическая адаптация). Предпосылкой к этому механизму служит внутрипопуляционная вариабельность уровня устойчивости к тому или иному фактору или группе факторов. Около трети поверхности суши испытывает дефицит влаги (годовое количество осадков 250 — 500 мм), а половина этой площади крайне засушлива (годовые осадки ниже 250 мм и испаряемость более 1000 мм). Для развития растений существенно, чтобы осадки относительно равномерно распределялись во время периодов активного роста. Однако в районах неустойчивого увлажнения часто бывают засушливые периоды именно в летние месяцы.
Засуха возникает как результат достаточно длительного отсутствия дождей, сопровождается высокой температурой воздуха и солнечной инсоляцией. Чаще она начинается с атмосферной засухи, характеризующейся низкой относительной влажностью воздуха. При длительном отсутствии дождей к атмосферной засухе добавляется почвенная засуха в связи с уменьшением (исчезновением) доступной для растений воды в почве. Во время суховея (атмосферная засуха, сопровождаемая сильным ветром) почвенная засуха может не возникать. В условиях засухи растения испытывают значительный водный дефицит.
Влияние недостатка воды на растение.Недостаток воды в тканях растений создается, когда расход воды при транспирации превышает ее поступление. Водный дефицит может возникнуть в жаркую солнечную погоду к середине дня, при этом увеличивается сосущая сила листьев, что активирует поступление воды из почвы. Растения регулируют уровень водного дефицита, меняя отверстость устьиц. Обычно при завядании листьев водный дефицит их восстанавливается в вечерние и ночные часы (временное завядание). Глубокое завядание наблюдается при отсутствии в почве доступной для растения воды. Это завядание чаще всего приводит растения к гибели.
Характерный признак устойчивого водного дефицита — сохранение его в тканях утром, а также прекращение выделения пасоки из срезанного стебля. Действие засухи в первую очередь приводит к уменьшению в клетках свободной воды, что изменяет гидратные оболочки белков цитоплазмы и сказывается на функционировании белков-ферментов. При длительном завядании снижается активность ферментов синтеза и активируются гидролитические процессы, в частности протеолиз, что ведет к увеличению содержания в клетках низкомолекулярных белков. В результате гидролиза полисахаридов в тканях накапливаются растворимые углеводы, отток которых из листьев замедлен. Под влиянием засухи в листьях снижается количество РНК вследствие уменьшения ее синтеза и активации ри-бонуклеаз. В цитоплазме наблюдается распад полирибосомных комплексов. Изменения, касающиеся ДНК, происходят лишь при длительной засухе. Из-за уменьшения свободной воды возрастает концентрация вакуолярного сока. Изменяется ионный состав клеток, облегчаются процессы выхода из них ионов.
В большинстве случаев суммарный фотосинтез при недостатке влаги снижается (см. рис. 4.16), хотя иногда на начальных этапах обезвоживания наблюдается некоторое увеличение его интенсивности. Снижение скорости фотосинтеза может быть следствием: 1) недостатка СО2 из-за закрывания устьиц, 2) нарушения синтеза хлорофиллов, 3) разобщения транспорта электронов и фотофосфорилирования, 4) изменений в фотохимических реакциях и реакциях восстановления СО2, 5) нарушения структуры хлоропластов, 6) задержки оттока ас-симилятов из листьев при длительном водном дефиците.


При обезвоживании у растений, не приспособленных к засухе, значительно усиливается интенсивность дыхания (возможно, из-за большого количества субстратов дыхания — Сахаров), а затем постепенно снижается. У засухоустойчивых растений в этих условиях существенных изменений дыхания не наблюдается или отмечается небольшое усиление.
В условиях водного дефицита быстро тормозятся клеточное деление и особенно растяжение, что приводит к формированию мелких клеток. Вследствие этого задерживается рост самого растения, особенно листьев и стеблей. Рост корней в начале засухи даже ускоряется и снижается лишь при длительном недостатке воды в почве. Корни реагируют на засуху рядом защитных приспособлений: опробковением, суберинизацией экзодермы, ускорением дифференцировки клеток, выходящих из меристемы, и др.
Таким образом, недостаток влаги вызывает значительные и постепенно усиливающиеся изменения большинства физиологических процессов у растений.
Влияние перегрева на физиологические процессы.Во время засухи наряду с обезвоживанием происходит перегрев растений. При действии высоких температур (35 С и  выше) наблюдаются два типа изменения вязкости цитоплазмы: чаще увеличение, реже снижение. Возрастание вязкости цитоплазмы замедляет ее движение (рис. 14.2), но процесс обратим даже при 5- минутном воздействии температуры 51°С. Высокая температура увеличивает концентрацию клеточного сока и проницаемость клеток для мочевины, глицерина, эозина и других соединений. В результате экзоосмоса веществ, растворенных в клеточном соке, постепенно снижается осмотическое давление. Однако при температурах выше 35°С вновь отмечается рост осмотического давления из-за усиления гидролиза крахмала и увеличения содержания моносахаров. Как следует из рис. 14.2, у листьев традесканции выход электролитов индуцируется под влиянием температуры более высокой по сравнению с температурой, меняющей вязкость цитоплазмы и ее движение. При этом потеря свойства полупроницаемости тонопласта (оцениваемая по выходу антоциана) вызывается лишь кратковременным действием очень высоких температур (57 — 64 °С).
выше) наблюдаются два типа изменения вязкости цитоплазмы: чаще увеличение, реже снижение. Возрастание вязкости цитоплазмы замедляет ее движение (рис. 14.2), но процесс обратим даже при 5- минутном воздействии температуры 51°С. Высокая температура увеличивает концентрацию клеточного сока и проницаемость клеток для мочевины, глицерина, эозина и других соединений. В результате экзоосмоса веществ, растворенных в клеточном соке, постепенно снижается осмотическое давление. Однако при температурах выше 35°С вновь отмечается рост осмотического давления из-за усиления гидролиза крахмала и увеличения содержания моносахаров. Как следует из рис. 14.2, у листьев традесканции выход электролитов индуцируется под влиянием температуры более высокой по сравнению с температурой, меняющей вязкость цитоплазмы и ее движение. При этом потеря свойства полупроницаемости тонопласта (оцениваемая по выходу антоциана) вызывается лишь кратковременным действием очень высоких температур (57 — 64 °С).
Процесс фотосинтеза более чувствителен к действию высоких температур, чем дыхание (см. рис. 4.14 и 14.2). Гидролиз полимеров, в частности белков, ускоряющийся при водном дефиците, значительно активируется при высокотемпературном стрессе. Распад белков идет с образованием аммиака, который может оказывать отравляющее действие на клетки у неустойчивых к перегреву растений. У жаростойких растений наблюдается увеличение содержания органических кислот, связывающих избыточный аммиак. Еще одним способом защиты от перегрева может служить усиленная транспирация, обеспечиваемая мощной корневой системой. В других случаях (суккуленты) жаростойкость определяется высокой вязкостью цитоплазмы и повышенным содержанием прочно связанной воды. При действии высоких температур в клетках растений индуцируется синтез стрессовых белков (белков теплового шока).
В сельскохозяйственной практике для повышения жароустойчивости растений применяют внекорневую обработку 0,05%-ным раствором солей цинка.
Приспособление растений к засухе.Как уже отмечалось, неблагоприятное действие засухи состоит в том, что растения испытывают недостаток воды или комплексное влияние обезвоживания и перегрева. У растений засушливых местообитаний — ксерофитов — выработались приспособления, позволяющие переносить периоды засухи.
Растения используют три основных способа защиты: 1) предотвращение излишней потери воды клетками (избегание высыхания), 2) перенесение высыхания, 3) избегание периода засухи. Наиболее общими являются приспособления для сохранения воды в клетках.
Группа ксерофитов очень разнородна. По способности переносить условия засухи различают следующие их типы (по П. А. Генкелю):
1. Суккуленты (по Н. А. Максимову — ложные ксерофиты) — растения, запасающие влагу (кактусы, алоэ, очиток, молодило, молочай). Вода концентрируется в листьях или стеблях, покрытых толстой кутикулой, волосками. Транспирация, фотосинтез и рост осуществляются медленно. Они плохо переносят обезвоживание. Корневая система распространяется широко, но на небольшую глубину.
2. Несуккулентные виды по уровню транспирации делятся на несколько групп.
а) Настоящие ксерофиты (эвксерофиты — полынь, вероника беловойлочная и др.). Растения с небольшими листьями, часто опушенными, жароустойчивы, транспирация невысокая, способны выносить сильное обезвоживание, в клетках высокое осмотическое давление. Корневая система сильно разветвлена, но на небольшой глубине.
б) Полуксерофиты (гемиксерофиты — шалфей, резак и др.). Обладают интенсивной транспирацией, которая поддерживается деятельностью глубокой корневой системы, часто достигающей грунтовых вод. Плохо переносят обезвоживание и атмосферную засуху. Вязкость цитоплазмы у них невелика.
в) Стипаксерофиты — степные злаки (ковыль и др.). Приспособлены к перенесению перегрева, быстро используют влагу летних дождей, но переносят лишь кратковременный недостаток воды в почве.
г) Пойкилоксерофиты (лишайники и др.) не способны регулировать свой водный режим и при значительном обезвоживании впадают в состояние покоя (анабиоз). Способны переносить высыхание.
3. Э ф е м е р ы — растения с коротким вегетационным периодом, совпадающим с периодом дождей (способ избегания засухи в засушливых местообитаниях).
Изучая физиологическую природу засухоустойчивости ксерофитов, Н. А. Максимов (1953) показал, что эти растения не являются сухолюбивыми: обилие воды в почве способствует их интенсивному росту. Устойчивость к засухе заключается в их способности переносить потерю воды.
Растения-мезофиты также могут приспосабливаться к засухе. Изучение приспособлений листьев к затрудненным условиям водоснабжения (В. Р. Заленский, 1904) показало, что анатомическая структура листьев различных ярусов на одном и том же растении зависит от уровня водоснабжения, освещенности и т. д. Чем выше по стеблю расположен лист, тем мельче его клетки, больше устьиц на единицу поверхности, а размер их меньше, гуще сеть проводящих пучков, сильнее развита па-лисадная паренхима и т. д. Такого рода закономерности изменений листового аппарата получили название закона Заленского. Было выяснено, что более высоко расположенные листья часто попадают в условия худшего водоснабжения (особенно у высоких растений), но обладают более интенсивной транспирацией. Устьица у листьев верхних ярусов даже при водном дефиците дольше остаются открытыми. Это, с одной стороны, поддерживает процесс фотосинтеза, а с другой — способствует увеличению концентрации клеточного сока, что позволяет им оттягивать воду от ниже расположенных листьев. Поскольку сходные особенности строения свойственны ряду ксерофитов, такая структура листьев получила название ксероморфной. Следовательно, возникновение ксероморфной структуры листьев — одно из анатомических приспособлений к недостатку воды, так же как заглубление устьиц в ткани листа, опушенность, толстая кутикула, редукция листьев и др.
Биохимические механизмы защиты предотвращают обезвоживание клетки, обеспечивают детоксикацию продуктов распада, способствуют восстановлению нарушенных структур цитоплазмы. Высокую водоудерживающую способность цитоплазмы в условиях засухи поддерживает накопление низкомолекулярных гидрофильных белков, связывающих в виде гидратных оболочек значительные количества воды. Этому помогает также взаимодействие белков с пролином, концентрация которого значительно возрастает в условиях водного стресса (см. 14.2), а также увеличение в цитоплазме содержания моносахаров.
 Интересным приспособлением, уменьшающим потерю воды через устьица, обладают суккуленты. Благодаря особенностям процесса фотосинтеза (САМ-метаболизм, см. 3.4.3) в дневные часы в условиях высокой температуры и сухости воздуха пустыни их устьица закрыты, поскольку СО2 фиксируется ночью.
Интересным приспособлением, уменьшающим потерю воды через устьица, обладают суккуленты. Благодаря особенностям процесса фотосинтеза (САМ-метаболизм, см. 3.4.3) в дневные часы в условиях высокой температуры и сухости воздуха пустыни их устьица закрыты, поскольку СО2 фиксируется ночью.
Детоксикация избытка образующегося при протеолизе аммиака осуществляется с участием органических кислот, количество которых возрастает в тканях при водном дефиците и высокой температуре. Процессы восстановления после прекращения действия засухи идут успешно, если сохранены от повреждения при недостатке воды и перегреве генетические системы клеток. Защита ДНК от действия засухи состоит в частичном выведении молекулы из активного состояния с помощью ядерных белков и, возможно, как в случае теплового стресса, с участием специальных стрессовых белков. Поэтому изменения количества ДНК обнаруживаются лишь при сильной длительной засухе.
Засуха вызывает существенные перестройки в гормональной системе растений: уменьшается содержание гормонов-активаторов роста — ауксина, цитокинина, гиббереллинов, стимуляторов роста фенольной природы и возрастает уровень абсцизо-вой кислоты и этилена. Приспособительный характер такого перераспределения очевиден, так как для поддержания роста необходима вода. Поэтому в условиях засухи от быстроты остановки ростовых процессов часто зависит выживание растения. При этом на ранних этапах засухи, по-видимому, главную роль играет стремительное возрастание содержания ингибиторов роста, поскольку даже в условиях сбалансированного водоснабжения клеток срочные реакции закрывания устьиц у растений осуществляются за счет ускоренного (в течение нескольких минут при водном дефиците 0,2 МПа) увеличения содержания АБК (см. 13.6.1). Пороговые величины водного потенциала, вызывающие увеличение АБК у растений-мезофитов, могут зависеть от степени засухоустойчивости растений: для кукурузы это 0,8 МПа, для сорго — 1 МПа. Содержание гормона в тканях в среднем увеличивается на порядок со скоростью 0,15 мкг/г сырой массы в час. Закрывание устьиц уменьшает потерю воды через транспирацию. Кроме того, АБК способствует запасанию гидратной воды в клетке, поскольку активирует синтез пролина, увеличивающего оводненность белков клетки в условиях засухи. АБК тормозит также синтез РНК, и белков, накапливаясь в корнях, задерживает синтез цитокинина. Таким образом, увеличение содержания АБК при водном дефиците уменьшает потерю воды через устьица, способствует запасанию гидратной воды белками и переводит обмен веществ клеток в режим «покоя».
В условиях водного стресса отмечается значительное выделение этилена. Так, в листьях пшеницы при уменьшении содержания воды на, 9% образование этилена возрастает в 30 раз в течение 4 ч. Выяснено, что водный стресс увеличивает активность синтетазы 1-аминоциклопропанкарбоновой кислоты, катализирующей ключевую реакцию биосинтеза этилена. При улучшении водного режима выделение этилена возвращается к норме. У многих растений при действии засухи (воздушной и почвенной) обнаружено также накопление ингибиторов роста фенольной природы (хлорогеновой кислоты, флавоноидов, фе-нолкарбоновых кислот).
Отмеченные выше изменения содержания фитогормонов-ин-гибиторов наблюдаются у растений-мезофитов при засухе. У пойкилоксерофитов, переходящих при наступлении засухи в состояние анабиоза, прекращение роста не связано с накоплением ингибиторов роста.
Снижение содержания гормонов-активаторов роста, в частности ПУК, происходит, по-видимому, вслед за остановкой роста. Например, в листьях подсолнечника, в верхушках стеблей и колосках пшеницы и других растений рост начинает подавляться уже при влажности почвы, составляющей 60% от полевой влагоемкости, а количество ауксинов заметно снижается в условиях почвенной засухи при влажности почвы около 30% от полевой влагоемкости. Уменьшение ауксина в тканях при засухе может быть связано с низким содержанием его предшественника — триптофана, а также с подавлением транспорта ауксинов по растению. Обработка растений в условиях засухи растворами ауксина, цитокинина, гиббереллина усугубляет отрицательное действие засухи. Однако опрыскивание растений цитокинином в период восстановления после засухи значительно улучшает состояние растений. Кроме того, цитокинин увеличивает жаростойкость растений (в частности улучшает всхожесть семян). Как предполагает О. Н. Кулаева (1973), это защитное действие цитокининов может быть связано с их влиянием на структурное и функциональное состояние макромолекулярных компонентов клетки, в частности на мембранные системы.
 Засухоустойчивость сельскохозяйственных растений повы шается в результате предпосевного закаливания (П. А. Генкель, 1934). Адаптация к обезвоживанию происходит в семенах, которые перед посевом после однократного намачивания вновь высушиваются. Для растений, выращенных из таких семян, характерны морфологические признаки ксероморфности, коррелирующие с их большей засухоустойчивостью. Растения различных мест обитания имеют и неодинаковые границы повреждающих низких температур. Так, растения Крайнего Севера без особого вреда зимой переносят охлаждение до - 60 °С. Такие центральноевропейские виды, как цветущие зимой маргаритка (Bellis perennis) или звездчатка (Stellaria media), могут перенести замерзание, а при повышении темпера туры продолжают свою жизнедеятельность. В то же время большинство теплолюбивых растений южного происхождения плохо переносит низкие положительные температуры (от 10°С и ниже). Например, растения какао погибают при 8 °С, хлоп чатник гибнет в течение суток при температуре от 1 до 3°С, прорастание зерновок и рост проростка кукурузы тормозится температурами почвы ниже 10 °С и т. д.
Засухоустойчивость сельскохозяйственных растений повы шается в результате предпосевного закаливания (П. А. Генкель, 1934). Адаптация к обезвоживанию происходит в семенах, которые перед посевом после однократного намачивания вновь высушиваются. Для растений, выращенных из таких семян, характерны морфологические признаки ксероморфности, коррелирующие с их большей засухоустойчивостью. Растения различных мест обитания имеют и неодинаковые границы повреждающих низких температур. Так, растения Крайнего Севера без особого вреда зимой переносят охлаждение до - 60 °С. Такие центральноевропейские виды, как цветущие зимой маргаритка (Bellis perennis) или звездчатка (Stellaria media), могут перенести замерзание, а при повышении темпера туры продолжают свою жизнедеятельность. В то же время большинство теплолюбивых растений южного происхождения плохо переносит низкие положительные температуры (от 10°С и ниже). Например, растения какао погибают при 8 °С, хлоп чатник гибнет в течение суток при температуре от 1 до 3°С, прорастание зерновок и рост проростка кукурузы тормозится температурами почвы ниже 10 °С и т. д.
Поэтому устойчивость растений к низким температурам подразделяют на холодостойкость, или устойчивость теплолюбивых растений к низким положительным температурам, и морозоустойчивость, или способность растений переносить температуры ниже 0°С.
 Холодостойкость.При помещении теплолюбивых растений в условия низкой положительной температуры отмечается постепенная потеря тургора клетками надземной части (например, листья огурца теряют тургор при 3°С на 3-й день, растение завядает и гибнет). Следовательно, при низких температурах может нарушаться доставка воды к транспирирующим органам.
Холодостойкость.При помещении теплолюбивых растений в условия низкой положительной температуры отмечается постепенная потеря тургора клетками надземной части (например, листья огурца теряют тургор при 3°С на 3-й день, растение завядает и гибнет). Следовательно, при низких температурах может нарушаться доставка воды к транспирирующим органам.
Однако при низкой температуре растение может погибнуть и в условиях, устраняющих влияние транспирапии, например, в пространстве, насыщенном парами воды. При этом проявляется «чистый» эффект пониженных температур на обмен веществ растений. У ряда видов наблюдается усиление распада белков и накопление в тканях растворимых форм азота. Основной причиной повреждающего действия низкой положительной температуры на теплолюбивые растения является нарушение функциональной активности мембран из-за перехода насыщенных жирных кислот, входящих в их состав, из жидко-кристаллического состояния в состояние геля при низкой температуре. Это приводит к неблагоприятным сдвигам в обмене веществ, а при длительном действии низкой температуры — к гибели растения.
Холодостойкость теплолюбивых сельскохозяйственных растений можно усилить предпосевным закаливанием семян. Наклюнувшиеся семена теплолюбивых культур (огурцы, томаты, дыня и др.) в течение нескольких суток выдерживают в чередующихся (через 12 ч) условиях низких положительных (1—5°С) и более высоких температур (10 —20 С). Таким же способом можно затем закалять рассаду. Холодостойкость повышается также при замачивании семян в 0,25%-ных растворах микроэлементов или нитрата аммония (в течение 20 ч для хлопчатника).
Морозоустойчивость.Морозы со средним годовым минимумом температуры воздуха ниже — 20 °С обычны на 42 % территории Земли. Поэтому очевидно значение понимания механизмов морозоустойчивости растений для сельскохозяйственного производства этих районов. В нашей стране большой вклад в изучение проблемы морозоустойчивости внесли работы Н. А. Максимова, И. И. Туманова и многих других исследователей. Быстрое понижение температуры в экспериментальных условиях сопровождается образованием льда внутри клеток и, как правило, их гибелью. Постепенное снижение температуры (0,5—ГС/ч), что обычно в естественных условиях, приводит к образованию льда в межклетниках. При этом образующиеся кристаллы льда вытесняют из межклетников воздух, и замерзшая ткань выглядит прозрачной. При оттаивании межклетники заполняются водой, которая затем поглощается клетками, если они не погибли от мороза.
 Основными причинами гибели клеток при низких отрицательных температурах являются: 1) их обезвоживание и 2) механическое сжатие льдом, повреждающее клеточные структуры.
Основными причинами гибели клеток при низких отрицательных температурах являются: 1) их обезвоживание и 2) механическое сжатие льдом, повреждающее клеточные структуры.
Обезвоживание возникает из-за оттягивания воды из клеток образующимися в межклетниках кристаллами льда. Это иссушающее действие льда, особенно при длительном действии низких температур, сходно с обезвоживанием, происходящим при засухе за счет испарения. При длительном действии мороза кристаллы льда вырастают до значительных размеров и, помимо сжатия клеток, могут повреждать плазмалемму.
Наиболее распространенные признаки повреждения от замерзания — потеря клетками тургора, инфильтрация межклетников водой и вымывание иомов из клеток. С помощью меченной тритием воды было показано, что при этом проницаемость клеточных мембран для воды не изменяется, что свидетельствует об отсутствии значительных сдвигов в липид-ной фазе мембран. Выход Сахаров и ионов К+ из клеток, по-видимому, связан с повреждением мембранных систем их активного транспорта (на основе АТРаз).
Приспособления растений к перенесению низких температур.Морозоустойчивые растения способны предотвращать или уменьшать действие низких отрицательных температур. Такие растения обладают приспособлениями, уменьшающими обезвоживание клетки.
1. Для предотвращения образования внутриклеточного льда при заморозках первостепенное значение имеет возможность быстрого транспорта свободной воды из клетки к местам внеклеточного образования льда, т. е. поддержание высокой проницаемости мембран в этих условиях. Такая возможность обеспечивается особенностями липидного состава мембран устойчивых растений. Общая реакция растений на низкие температуры — увеличение в составе мембран количества ненасыщенных жирных кислот. Это обусловливает снижение температуры фазового перехода липидов из жидко-кристаллического состояния в гель до величины, лежащей ниже точки замерзания у морозостойких растений, а у неустойчивых растений она выше 0°С. Фазовые переходы мембран из жидко-кристаллического в твердое (гель) состояние на 1/3 снижают проницаемость липидных мембран. Поэтому понижение температуры фазового перехода липидов у морозоустойчивых растений сохраняет высокую проницаемость мембран при замораживании.
2. Перенесению морозов способствует также усиление процессов синтеза веществ,защищающих ткани (криопротекторов). К ним относятся прежде всего полимеры, способные связывать значительные количества воды, — гидрофильные белки, моно-и олигосахариды. Вода, связываемая в виде гидратных оболочек этими молекулами, не замерзает и не транспортируется, оставаясь в клетке. Таким образом клетки защищаются от внутриклеточного льда и чрезмерного обезвоживания. У морозоустойчивых растений при действии низких температур усиливается гидролиз крахмала и в цитоплазме накапливаются сахара, у большинства растений возрастает синтез водорастворимых белков. Чем выше их содержание, тем больше способность клетки к выживанию в условиях низких температур.
 Другой тип полимеров-криопротекторов — молекулы геми-целлюлоз (ксиланы, арабиноксштаны), выделяемые в клеточную стенку. Они обволакивают кристаллы льда и тормозят их рост. В итоге образуются более мелкие кристаллы, меньше повреждающие клетку.
Другой тип полимеров-криопротекторов — молекулы геми-целлюлоз (ксиланы, арабиноксштаны), выделяемые в клеточную стенку. Они обволакивают кристаллы льда и тормозят их рост. В итоге образуются более мелкие кристаллы, меньше повреждающие клетку.
3. У морозоустойчивых растений в период подготовки к зиме накапливаются запасные вещества, которые могут использоваться затем при возобновлении роста. Существенна также устойчивость их к болезням, опасность возникновения которых возрастает при повреждении тканей морозом.
Холодоустойчивость растений можно повысить с помощью закалки. Закаливание подготавливает весь комплекс защитных средств растения, о котором говорилось выше. Закалка ускоряется при остановке ростовых процессов у растения и осуществляется при постепенном снижении температуры, а для ряда объектов — и при укороченном фотопериоде.
 В нашей стране теория закаливания к низким температурам разработана И. И. Тумановым. Согласно этой теории растения для приобретения свойства морозостойкости должны пройти три этапа подготовки: переход в состояние покоя, затем первую и вторую фазы закаливания. Вступление в состояние покоя без последующих этапов лишь немного повышает морозоустойчивость. Переход в состояние покоя сопровождается смещением баланса фитогормонов в сторону уменьшения содержания ауксина и гиббереллинов и увеличения абсцизовой кислоты. Обработка растений в этот период ингибиторами роста (хлорхолинхлоридом или трийодбензойной кислотой) повышает устойчивость растений к низким температурам, а обработка ИУК или ГА — понижает ее. У древесных растений покой наступает в начале осени и в первую фазу закаливания лишь углубляется; у травянистых — переход в состояние покоя сопровождает первую фазу закаливания.
В нашей стране теория закаливания к низким температурам разработана И. И. Тумановым. Согласно этой теории растения для приобретения свойства морозостойкости должны пройти три этапа подготовки: переход в состояние покоя, затем первую и вторую фазы закаливания. Вступление в состояние покоя без последующих этапов лишь немного повышает морозоустойчивость. Переход в состояние покоя сопровождается смещением баланса фитогормонов в сторону уменьшения содержания ауксина и гиббереллинов и увеличения абсцизовой кислоты. Обработка растений в этот период ингибиторами роста (хлорхолинхлоридом или трийодбензойной кислотой) повышает устойчивость растений к низким температурам, а обработка ИУК или ГА — понижает ее. У древесных растений покой наступает в начале осени и в первую фазу закаливания лишь углубляется; у травянистых — переход в состояние покоя сопровождает первую фазу закаливания.
В течение первой фазы закаливания (озимые злаки проходят первую фазу на свету при 0,5 —2°С за 6 — 9 дней; древесные — за 30 дней) при пониженных положительных температурах (до О °С) останавливается рост (если растения не находятся в состоянии покоя), в клетках накапливаются соединения, выполняющие защитную функцию (сахара, растворимые белки и др.), в мембранах возрастает содержание ненасыщенных жирных кислот, снижается точка замерзания цитоплазмы, отмечается некоторое уменьшение внутриклеточной воды, что тормозит образование внутриклеточного льда.
В период прохождения второй фазы закаливания (постепенное понижение температуры до — 10, — 20 °С и ниже со скоростью 2 —3°С в сутки) в межклетниках образуется лед и начинают функционировать механизмы защиты от обезвоживания, подготовленные в течение первой фазы.
 На морозоустойчивость, как и на холодостойкость растений, положительное влияние оказывают микроэлементы. Так, цинк повышает содержание связанной воды и усиливает накопление Сахаров, а молибден способствует увеличению содержания общего и белкового азота. Сходный эффект оказывают кобальт, медь, ванадий и др.
На морозоустойчивость, как и на холодостойкость растений, положительное влияние оказывают микроэлементы. Так, цинк повышает содержание связанной воды и усиливает накопление Сахаров, а молибден способствует увеличению содержания общего и белкового азота. Сходный эффект оказывают кобальт, медь, ванадий и др.
Изучение солеустойчивости растений имеет большое практическое значение, поскольку океаны, воды которых содержат 3 — 4% солей, занимают около 75% поверхности Земли, 25% почв планеты засолены, а треть земель в мире, подвергшихся ирригации, уже изменены в сторону избытка солей вследствие плохого дренажа.
Согласно Б. П. Строганову (1962), по степени засоления различают практически незасоленные, слабозасоленные, сред-незасоленные почвы и солончаки. Тип засоления определяется по содержанию анионов в почве: хлоридное, сульфатное, суль-фатно-хлоридное, хлоридно-сульфатные и карбонатное. Преобладающим катионом в таких почвах является натрий, но встречаются также карбонатно-магниевое и хлоридно-магние-вое засоление.
Растения, приспособленные к существованию в условиях избыточного засоления, называют галофитами (от греч. «galos» - соль, «phyton» - растение). Они отличаются от гликофи-тов — растений незасоленных водоемов и почв — рядом анатомических особенностей и особенностей обмена веществ. Галофиты защищаются от избыточной концентрации солей тремя основными способами: 1) поглощением большого количества солей и концентрированием их в вакуолярном соке, что приводит к созданию высокого осмотического давления; 2) выведением поглощаемых солей из клеток вместе с водой с помощью специализированных солевых желёзок и удалением избытка солей с опавшими листьями (см. 9.2.4); одна из разновидностей этого способа — локализация поглощаемых солей в вакуолях клеток-головок специализированных волосков, которые впоследствии обламываются (см. рис. 9.2); 3) ограниченным поглощением солей клетками корней.
Все галофиты можно разделить на три группы:
1. Настоящие галофиты (эвгалофиты) — наиболее солеустойчивые растения, накапливающие в вакуолях значительные концентрации солей. Растут на влажных засоленных почвах. Вследствие высокого осмотического давления в клетках растения обладают большой сосущей силой, позволяющей поглощать воду из сильно засоленной почвы. Для растений этой группы характерна мясистость листьев (галосуккулентность), которая исчезает при выращивании их на незасоленных почвах. Типичные представители настоящих галофитов — солерос.(Salicornia herbaced) и сведа (Suaeda maritima).
 2. Солевыделяющие галофиты (криногалофиты), поглощая соли, не накапливают их внутри тканей, а выводят из клеток с помощью секреторных желёзок, расположенных на листьях. Выделение солей желёзками осуществляется с помощью ионных насосов и сопровождается транспортом болыцих количеств воды (см. 9.2.4). Соли оседают белыми налетами на листьях. Часть солей удаляется с опадающими листьями. Эти особенности характерны для кермека (Statice gmelini), тамарикса (Tamarix speciosa) и др. Галофиты этих двух групп называют также солянками. У некоторых видов (Atriplex) при выделении солей в вакуоль клетки-головки волоска (с последующим ее обламыванием и восстановлением) избавление от избытка солей происходит без поглощения больших количеств воды (см. рис. 9.2).
2. Солевыделяющие галофиты (криногалофиты), поглощая соли, не накапливают их внутри тканей, а выводят из клеток с помощью секреторных желёзок, расположенных на листьях. Выделение солей желёзками осуществляется с помощью ионных насосов и сопровождается транспортом болыцих количеств воды (см. 9.2.4). Соли оседают белыми налетами на листьях. Часть солей удаляется с опадающими листьями. Эти особенности характерны для кермека (Statice gmelini), тамарикса (Tamarix speciosa) и др. Галофиты этих двух групп называют также солянками. У некоторых видов (Atriplex) при выделении солей в вакуоль клетки-головки волоска (с последующим ее обламыванием и восстановлением) избавление от избытка солей происходит без поглощения больших количеств воды (см. рис. 9.2).
3. Соленепроницаемые галофиты (гликогалофиты) растут на менее засоленных почвах. Высокое осмотическое давление в их клетках поддерживается за счет продуктов фотосинтеза, а клетки малопроницаемы для солей. Типичные представители этой группы — полынь (Artemisia salina), различные виды кохии (Kochia).
Тип засоления почвы влияет на структуру органов растений. При хлоридном засолении растения приобретают суккулентные черты — мясистость листьев, развиваются водоносные ткани. При сульфатном засолении у растений преобладают черты ксе-роморфности. Соответственно в условиях хлоридного засоления невысока интенсивность дыхания и фотосинтеза, замедлено выделение воды; при сульфатном засолении эти показатели выше.
Растения-гликофиты в условиях засоления также обнаруживают определенную способность к перенесению избытка солей. Из сельскохозяйственных растений относительно солеустой-чивы ячмень, сахарная свекла, хлопчатник; мягкая пшеница устойчивее твердой. При длительном выращивании в условиях засоления для ячменя, проса, томатов выявлено значительное повышение их солеустойчивости без потери продуктивности. Значительный избыток солей в почве может привести гликофиты к гибели, особенно при засолении в фазы активного роста и цветения. Засоление сильнее всего подавляет рост клеток растяжением и в меньшей степени влияет на их деление. Избыточное засоление нарушает азотный обмен, что способствует накоплению промежуточных продуктов азотного обмена, особенно аминов и диаминов (кадаверин, путресцин) и аммиака. Эти соединения могут оказывать сильное токсическое действие на растения. Засоление приводит к возникновению дисбаланса между поглощением натрия, калия и магния; интенсивное поглощение Na уменьшает поступление К и Mg в клетки. Осмотическое давление почвенного раствора вызывает повреждения лишь при очень высоком засолении, когда оно затрудняет поглощение воды.
В сельскохозяйственном производстве основным методом борьбы с засолением является мелиорация засоленных почв, создание надежного дренажа и промывка почв после сбора урожая. На солонцах (почвы, содержащие много натрия) мелиорацию осуществляют с помощью гипсования, которое приводит к вытеснению натрия из почвенного поглощающего комплекса и замещению его кальцием.
 Внесение в почву микроэлементов улучшает ионный обмен растений в условиях засоления. Солеустойчивость растений увеличивается после применения предпосевного закаливания семян. Для семян хлопчатника, пшеницы, сахарной свеклы достаточна обработка в течение часа 3%-ным раствором NaCl с последующим промыванием водой (1,5 ч). Этот прием повышает устойчивость растений при хлоридном засолении. Для закалки к сульфатному засолению семена в течение суток вымачивают в 0,2%-ном растворе сульфата магния.
Внесение в почву микроэлементов улучшает ионный обмен растений в условиях засоления. Солеустойчивость растений увеличивается после применения предпосевного закаливания семян. Для семян хлопчатника, пшеницы, сахарной свеклы достаточна обработка в течение часа 3%-ным раствором NaCl с последующим промыванием водой (1,5 ч). Этот прием повышает устойчивость растений при хлоридном засолении. Для закалки к сульфатному засолению семена в течение суток вымачивают в 0,2%-ном растворе сульфата магния.
В настоящее время наряду с выведением солеустойчивых сортов культурных растений интенсивно развиваются методы генной инженерии, которые позволяют повысить устойчивость растений к неблагоприятным факторам. Задача заключается в том, чтобы найти комплекс генов, ответственный, в частности, за солеустойчивость, и научиться вводить их в клетки неустойчивых растений.
Условия кислородной недостаточности возникают при временном или постоянном переувлажнении, при заболачивании почвы, при вымокании растений, образовании ледяной корки на озимых посевах, в результате создания асфальтовых покрытий в городах, в процессе орошаемого землепользования, при хранении сельскохозяйственной продукции и т. п. Наиболее часто оказываются в этих условиях озимые хлеба (пшеница, рожь, ячмень), а также соя, рис, хлопчатник, некоторые древесные растения — ель, береза, сосна, ива, ольха.
Несмотря на низкое содержание кислорода в заболоченных или затопляемых почвах, многие виды растений приспособились к произрастанию на них. Как видно из схемы (Т. В. Чиркова, 1984), приспособления к существованию при недостатке кислорода направлены, с одной стороны, на сохранение близкого к нормальному уровню содержания кислорода в тканях, а с другой — на адаптацию к функционированию при пониженной концентрации кислорода в среде:
У растений, корни которых постоянно испытывают недостаток содержания кислорода, в процессе длительной эволюции появились разнообразные изменения в морфолого-анатомиче-ском строении тканей: разрастания основания стебля, образование дополнительной поверхностной корневой системы, вентиляционных систем межклетников, аэренхимы, необходимых для транспорта кислорода из надземных частей растения в корни. Эти приспособления способствуют тому, что растение получает возможность избежать действия неблагоприятного фактора. В обеспечении кислородом корневой системы, наиболее часто страдающей от недостатка атмосферного кислорода, существенна роль листьев, а у древесных — и чечевичек ветвей и стволов. У ряда растений доля кислорода, поступающего в корни за счет транспорта его из надземной части, составляет 8 — 25% от потребляемого в условиях нормальной аэрации.
Использование внешнего источника кислорода и доставка. его в корни, находящиеся в условиях гипоксии или аноксии (не- достатка или отсутствия кислорода), не может полностью обеспечить потребности растения, поскольку значительная часть О2 используется на дыхание в процессе транспортировки.

Поэтому появляется необходимость возникновения приспособлений к существованию растений при пониженной концентрации кислорода. Эти приспособления по своей сути являются физиолого-биохимическими (метаболическими). Из схемы видно, что они прежде всего связаны с процессом дыхания. При недостатке кислорода один тип приспособлений состоит в снижении общей интенсивности дыхания и использования дыхательных субстратов, хотя запасы субстратов у растений, характеризующихся такой реакцией (например, у болотных), могут быть больше, чем у мезофитов.
Другой тип приспособлений связан с перестройками в путях дыхания, необходимыми прежде всего для поддержания синтеза АТР, достаточного для сохранения жизнедеятельности растений. При гипоксии приспособительную роль может играть увеличение активности пентозофосфатного пути дыхания при условии, что есть возможность окисления образующегося в ходе его NADPH.
 При усилении анаэробиоза преобладающим становится гликолитический путь катаболизма глюкозы. Несмотря на малую энергетическую эффективность гликолиза, роль его для временного переживания условий анаэробиоза очень велика, особенно если функционируют и системы деток-сикации продуктов анаэробного распада (этилового спирта, молочной кислоты). Детоксикация состоит как в удалении этих веществ, так и во включении их в обмен. Отсутствие накопления продуктов брожений в токсических концентрациях, характерное для устойчивых к кислородному дефициту растений, способствует возобновлению и повышению активности гликолиза. Для функционирования гликолиза очень важно также окисление восстановленного NADH, образующегося в ходе этого процесса. У устойчивых растений, возможно, происходит обращение дикарбоновой части цикла Кребса — в малатдегидрогеназной реакции и в реакции, сопряженной с образованием янтарной кислоты.
При усилении анаэробиоза преобладающим становится гликолитический путь катаболизма глюкозы. Несмотря на малую энергетическую эффективность гликолиза, роль его для временного переживания условий анаэробиоза очень велика, особенно если функционируют и системы деток-сикации продуктов анаэробного распада (этилового спирта, молочной кислоты). Детоксикация состоит как в удалении этих веществ, так и во включении их в обмен. Отсутствие накопления продуктов брожений в токсических концентрациях, характерное для устойчивых к кислородному дефициту растений, способствует возобновлению и повышению активности гликолиза. Для функционирования гликолиза очень важно также окисление восстановленного NADH, образующегося в ходе этого процесса. У устойчивых растений, возможно, происходит обращение дикарбоновой части цикла Кребса — в малатдегидрогеназной реакции и в реакции, сопряженной с образованием янтарной кислоты.
При отсутствии в анаэробных условиях кислорода как конечного акцептора электронов приспособительными оказываются процессы так называемого токсического эндогенного окисления, в ходе которого электроны переносятся на такие вещества, как нитраты, двойные связи ненасыщенных соединений (жирные кислоты, каротиноиды). Перенос электронов на подобные акцепторы играет роль «запасного выхода» и способствует продлению жизни растений в описываемых условиях.
В результате указанных трансформаций обмена у устойчивых к недостатку кислорода растений обеспечивается длительная стабильность внутриклеточной кислотности и окислительно-восстановительного равновесия, тормозится распад углеводов, белков и липидов и даже сохраняется способность к синтезу некоторых белков-ферментов, играющих адаптивную роль. Таким образом, устойчивость к гипо- и аноксии характеризуется целым комплексом анатомо-морфологических и физиолого-биохимических приспособлений, обеспечивающих в данных условиях функционирование обмена веществ.
В настоящее время вопрос о повышении устойчивости растений к недостатку кислорода только начинает разрабатываться. Одним из агротехнических мероприятий, повышающих устойчивость растений к избыточному водоснабжению, является обработка растений и замачивание семян в растворах хлор-холинхлорида. Положительное влияние хлорхолинхлорида отражается на развитии генеративных органов и завязывании зерновок в колосьях пшеницы при ее затоплении. Уменьшение подавления роста и повышение урожая при затоплении ячменя наблюдалось также при предпосевном намачивании семян в 0,001—0,0001 %-ном растворе никотиновой кислоты. Близкие результаты были получены и при предпосевной обработке семян ячменя 0,1%-ным раствором сульфата марганца. В условиях гипо- и аноксии продлевают жизнедеятельность растений, способных к нитратному дыханию, подкормки нитратами.
 Газоустойчивость — это способность растений сохранять жизнедеятельность при действии вредных газов. На степень газоустойчивости растений влияют физико-географические и метеорологические условия. Растения не обладают сформировавшейся в ходе эволюции системой адаптации к вредным газам, и поэтому способность противостоять повреждающему действию газов основывается на механизмах устойчивости их к другим неблагоприятным факторам. Это связано с тем, что современная нам флора формировалась в условиях, при которых содержание вредных газов (вследствие вулканической деятельности, пожаров, химических процессов) в атмосферном воздухе было очень мало. Этот состав воздуха сформировался около 1 млрд. лет назад как следствие жизнедеятельности автотрофов. По-видимому, освобождение первичной атмосферы Земли от аммиака, сероводорода, метана, оксида углерода и др. активно осуществляли растения-автотрофы протерозойской и палеозойской эр, которые должны были обладать механизмами газоустойчивости. Но по мере увеличения кислорода в атмосферном воздухе и уменьшения содержания в нем вредных газов эти механизмы были утрачены.
Газоустойчивость — это способность растений сохранять жизнедеятельность при действии вредных газов. На степень газоустойчивости растений влияют физико-географические и метеорологические условия. Растения не обладают сформировавшейся в ходе эволюции системой адаптации к вредным газам, и поэтому способность противостоять повреждающему действию газов основывается на механизмах устойчивости их к другим неблагоприятным факторам. Это связано с тем, что современная нам флора формировалась в условиях, при которых содержание вредных газов (вследствие вулканической деятельности, пожаров, химических процессов) в атмосферном воздухе было очень мало. Этот состав воздуха сформировался около 1 млрд. лет назад как следствие жизнедеятельности автотрофов. По-видимому, освобождение первичной атмосферы Земли от аммиака, сероводорода, метана, оксида углерода и др. активно осуществляли растения-автотрофы протерозойской и палеозойской эр, которые должны были обладать механизмами газоустойчивости. Но по мере увеличения кислорода в атмосферном воздухе и уменьшения содержания в нем вредных газов эти механизмы были утрачены.
Загрязнение атмосферы, связанное с расширением производственной деятельности человека, возрастает в таких катастрофических масштабах, что системы авторегуляции биосферы уже не справляются с его очисткой. В результате различных видов деятельности человека (промышленность, автотранспорт и др.) в воздух выделяются более 200 различных компонентов. К ним относятся газообразные соединения: сернистый газ (SO2), оксиды азота (NO, NO2), угарный газ (СО), соединения фтора и др., углеводороды, пары кислот (серной, сернистой, азотной, соляной), фенола и др., твердые частицы сажи, золы, пыли, содержащие токсические оксиды свинца, селена, цинка и т. д. В промышленно развитых странах на 52,6% воздух загрязнен деятельностью транспорта, на 18,1% —отопительными системами, на 17,9% — промышленными процессами и на 1,9 и 9,5% — за счет сжигания мусора и других процессов соответственно.
Загрязняющие атмосферный воздух компоненты (эксгалаты) по величине частиц, скорости оседания под действием силы тяжести и электромагнитному спектру подразделяют на пыль, пары, туманы и дым.
Газы и пары, легко проникая в ткани растений через устьица, могут непосредственно влиять на обмен веществ клеток, вступая в химические взаимодействия уже на уровне клеточных стенок и мембран. Пыль, оседая на поверхности растения, закупоривает устьица, что ухудшает газообмен листьев, затрудняет поглощение света, нарушает водный режим.
По убыванию токсичности действия на растения газы можно расположить в следующие ряды: 1) F2 > С12 > SO2 > NO > >СО> СО2 или 2) С12 > SO2 > NH3 > HCN > H2S.
Кислые газы и пары более токсичны для растений, чем для животных, для которых второй ряд газов выглядит следующим образом: HCN > H2S > С12 > SO2 > NH3.
 Наиболее сильно газы воздействуют на процессы в листьях. Косвенный эффект загрязнения атмосферы проявляется через почву, где газы влияют на микрофлору, почвенный поглощающий комплекс и корни растений. Кислые газы и кислые дожди нарушают водный режим тканей, приводят к постоянному за-кислению цитоплазмы клеток, изменению работы транспортных систем мембран (плазмалеммы, хлоропластов), накоплению Са, Zn, Pb, Сu. В этих условиях интенсивность фотосинтеза снижается из-за нарушения мембран хлоропластов. Кроме того, на свету быстро разрушаются хлорофилл а и каротин, меньше — хлорофилл b и ксантофиллы. Особенно неблагоприятно на пигментную систему хлоропластов действуют SO2 и С12, аммиак же уменьшает содержание каротина и ксантофилла, мало влияя на хлорофиллы.
Наиболее сильно газы воздействуют на процессы в листьях. Косвенный эффект загрязнения атмосферы проявляется через почву, где газы влияют на микрофлору, почвенный поглощающий комплекс и корни растений. Кислые газы и кислые дожди нарушают водный режим тканей, приводят к постоянному за-кислению цитоплазмы клеток, изменению работы транспортных систем мембран (плазмалеммы, хлоропластов), накоплению Са, Zn, Pb, Сu. В этих условиях интенсивность фотосинтеза снижается из-за нарушения мембран хлоропластов. Кроме того, на свету быстро разрушаются хлорофилл а и каротин, меньше — хлорофилл b и ксантофиллы. Особенно неблагоприятно на пигментную систему хлоропластов действуют SO2 и С12, аммиак же уменьшает содержание каротина и ксантофилла, мало влияя на хлорофиллы.
Дыхание в условиях загрязнения, как правило, вначале возрастает, а затем снижается по мере развития повреждений. Все эти изменения нарушают рост растений, ускоряют процессы старения в них. Очень сильно страдают от кислых газов хвойные породы (суховершинность, ослабление роста стволов в толщину, уменьшение длины и увеличение числа хвоинок на побеге, быстрая потеря хвои). При длительном действии кислых газов наблюдаются значительные изменения в фитоце-нозах: утрата лесных пород, развитие сорной травянистой растительности. У лиственных пород кислые газы вызывают уменьшение размеров и количества листьев, индуцируют появление у них черт ксероморфности.
По характеру реакции у растений различают газочувствительность (т. е. скорость и степень проявления патологических процессов под влиянием газов) и газоустойчивость. Для газоустойчивости существенна способность растений 1) регулировать поступление токсичных газов, 2) поддерживать буферность цитоплазмы и ее ионный баланс, 3) осуществлять детоксикацию образующихся ядов. В итоге в условиях задымления это способствует поддержанию фотосинтеза и синтетических процессов на достаточно высоком уровне.
Регуляция поглощения газов определяется прежде всего чувствительностью устьиц к газам; под их влиянием (особенно SO2) газоустойчивые виды быстро закрывают устьица. Устойчивость к токсическим газам может быть связана и с уровнем в клетках катионов (К+ , Na+ , Ca2+), способных нейтрализовать ангидриды кислот. Обычно растения, устойчивые к засухе, засолению и другим стрессам, имеют и более высокую газоустойчивость, возможно, из-за способности регулировать водный режим и ионный состав. На это указывают усиление сернистым газом признаков ксероморфности листьев, а хлором — признаков суккулентности. Проверка газоустойчивости (по SO2) большого числа видов растений (В. С. Николаевский, 1979) позволила разделить их на три группы: устойчивые, сред-неустойчивые и неустойчивые. Наиболее устойчивые к SO2 древесные породы (вяз, жимолость, клен, лох) оказались устойчивыми также к хлору, фтору, диоксиду азота.
 Газоустойчивость растений повышается при оптимизации минерального питания и закалке семенного материала. Замачивание семян в слабых растворах соляной и серной кислот повышает устойчивость растений к кислым газам. Хотя загрязнение атмосферного воздуха наносит большой ущерб растительности, именно растения наряду с регуляцией водного, ветрового и других режимов среды представляют собой мощный фактор, очищающий атмосферу.
Газоустойчивость растений повышается при оптимизации минерального питания и закалке семенного материала. Замачивание семян в слабых растворах соляной и серной кислот повышает устойчивость растений к кислым газам. Хотя загрязнение атмосферного воздуха наносит большой ущерб растительности, именно растения наряду с регуляцией водного, ветрового и других режимов среды представляют собой мощный фактор, очищающий атмосферу.
в Биологический эффект ионизирующего излучения является результатом влияния радиации на многих уровнях — от моле кулярного до организменного и популяционного. Первичные механизмы действия всех типов излучения на живой организм сходны. Их общая особенность состоит в том, что значи тельный биологический эффект вызывается слабой энергией и небольшим числом первичных радиационно-химических ре акций. Например, при гамма-облучении дозой около 10 Гр (1000 р)1, летальной для млекопитающих, поглощается энер гия, равная 8,4 кДж/г, достаточная лишь для повышения тем пературы на 0,001°С.
Различают прямое и косвенное действие радиации на живые организмы. Прямое действие состоит в радиационно-химических превращениях молекул в месте поглощения энергии излучения. Прямое попадание в молекулу переводит ее в возбужденное или ионизированное состояние. Поражающее действие связано с ионизацией молекулы. Непрямое, или косвенное, действие радиации состоит в повреждениях молекул, мембран, органоидов, клеток, вызываемых продуктами радио-лиза воды, количество которых в клетке при облучении очень велико. Заряженная частица излучения, взаимодействуя с молекулой воды, вызывает ее ионизацию:
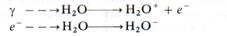
Ионы воды за время жизни 10-15 —10-10 с способны образовать химически активные свободные радикалы и перокси-ды:
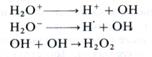
В присутствии растворенного в воде кислорода возникают также мощный окислитель НО2 (Н+ + О2→НО2) и новые перок-сиды (НО2+ Н→Н2О2) и т. д. Эти сильные окислители за время жизни 10-6-10-5 с могут повредить (изменить) многие биологически важные молекулы — нуклеиновые кислоты, белки-ферменты, липиды мембран и др. Кроме того, при взаимодействии радикалов воды с органическими веществами в присутствии кислорода образуются органические пероксиды, что также способствует лучевому повреждению молекул и структур клетки.
 При понижении концентрации кислорода в среде (ткани) уменьшается эффект лучевого поражения, а при увеличении его концентрации действие радиации усиливается. Этот «кислородный эффект» проявляется на всех уровнях организации биологических объектов — от молекулярного до тканевого.
При понижении концентрации кислорода в среде (ткани) уменьшается эффект лучевого поражения, а при увеличении его концентрации действие радиации усиливается. Этот «кислородный эффект» проявляется на всех уровнях организации биологических объектов — от молекулярного до тканевого.
Прямое действие радиации на молекулы объясняют теория «мишеней или попаданий» и вероятностная гипотеза. Согласно первой попадание ионизирующей частицы в чувствительную часть (мишень) молекулы или структуры клетки вызывает ее повреждение, генетические изменения и гибель. Обнаружено, что с увеличением дозы количество повреждений в облучаемом объеме увеличивается в геометрической прогрессии, причем повреждение может быть результатом как одного попадания, так и нескольких. По вероятностной гипотезе взаимодействие излучения с мишенью происходит по принципу случайности, а реакция на излучение зависит от состояния биологической системы в момент действия излучения.