СИСТЕМЫ РЕГУЛЯЦИИ И ИНТЕГРАЦИИ У РАСТЕНИЙ 1 страница
«...дифференциация и специализация неизбежно связаны с соподчинением частей, с интеграцией. Из этого ясно выступает возрастающее значение интегрирующих факторов в более сложных организмах».
И. И. Шмальгаузен
Сложное строение растительного организма, дифференцированного на большое количество специализированных органоидов, клеток, тканей и органов, требует и совершенных систем управления.
 Целостность всякого, в том числе и растительного, организма обеспечивается системами регуляции, управления и интеграции. В технике под регуляцией обычно понимают поддержание значений параметров системы в заданных границах. Управление — это процесс перевода системы из одного состояния в другое путем воздействия на ее переменные. Однако в более широком смысле термин «регуляция» включает в себя и процессы управления. В таком более широком смысле этот термин употребляется и в биологии. Регуляция обеспечивает гомеостаз организма, т. е. сохранение постоянства параметров внутренней среды, а также создает условия для его развития (эпигенеза). На всех уровнях организации гомеостаз обеспечивается отрицательными обратными связями, эпигенез — преимущественно положительными обратными связями.
Целостность всякого, в том числе и растительного, организма обеспечивается системами регуляции, управления и интеграции. В технике под регуляцией обычно понимают поддержание значений параметров системы в заданных границах. Управление — это процесс перевода системы из одного состояния в другое путем воздействия на ее переменные. Однако в более широком смысле термин «регуляция» включает в себя и процессы управления. В таком более широком смысле этот термин употребляется и в биологии. Регуляция обеспечивает гомеостаз организма, т. е. сохранение постоянства параметров внутренней среды, а также создает условия для его развития (эпигенеза). На всех уровнях организации гомеостаз обеспечивается отрицательными обратными связями, эпигенез — преимущественно положительными обратными связями.
 В ходе эволюции сначала должны были возникнуть внутриклеточные системы регуляции. К ним относятся регуляция на уровне ферментов, генетическая и мембранная регуляции (рис. 2.1,I). Все эти системы регуляции тесно связаны между собой. Например, свойства мембран зависят от генной активности, а дифференциальная активность самих генов находится под контролем мембран. Больше того, в основе всех форм внутриклеточной регуляции лежит единый первичный принцип, который можно назвать рецепторно-конфор-мационным. Во всех случаях белковая молекула — будь то фермент, рецептор или регуляторный белок - «узнает» специфический для нее фактор и, взаимодействуя с ним, изменяет свою конфигурацию. В мультикомпонентных комплексах ферментов, генов и мембран конформационные изменения молекул-рецепторов кооперативно передаются на весь комплекс, влияя на его функциональную активность.
В ходе эволюции сначала должны были возникнуть внутриклеточные системы регуляции. К ним относятся регуляция на уровне ферментов, генетическая и мембранная регуляции (рис. 2.1,I). Все эти системы регуляции тесно связаны между собой. Например, свойства мембран зависят от генной активности, а дифференциальная активность самих генов находится под контролем мембран. Больше того, в основе всех форм внутриклеточной регуляции лежит единый первичный принцип, который можно назвать рецепторно-конфор-мационным. Во всех случаях белковая молекула — будь то фермент, рецептор или регуляторный белок - «узнает» специфический для нее фактор и, взаимодействуя с ним, изменяет свою конфигурацию. В мультикомпонентных комплексах ферментов, генов и мембран конформационные изменения молекул-рецепторов кооперативно передаются на весь комплекс, влияя на его функциональную активность.
С появлением многоклеточных организмов развиваются и совершенствуются межклеточные системы регуляции. Они включают в себя по крайней мере трофическую, гормональную и электрофизиологическую системы (рис. 2.1,II). Такие взаимодействия сразу обнаруживаются при попытках культивировать те или иные части и органы растений в изолированном виде. Во всех случаях для поддержания жизни изолированных частей в инкубационную среду необходимо добавлять трофические и гормональные факторы, в норме поступающие из других органов целого растения.
 В последующих разделах внутриклеточные и межклеточныесистемы регуляции будут рассмотрены более детально. ' Изостерическая регуляция активности ферментов осуществляется на уровне их каталитических центров. Реакционная способность и направленность работы каталитического центра прежде всего зависят от количества субстрата (закон действия масс). Интенсивность работы фермента определяется также наличием коферментов (для двухкомпонентных энзимов), кофакторов (специфически действующих катионов) и активаторов или ингибиторов, действующих на уровне каталитического центра. Активность тех или других ферментов может быть связана с конкуренцией за общие субстраты и коферменты, что является одним из способов взаимодействия различных метаболических циклов.
В последующих разделах внутриклеточные и межклеточныесистемы регуляции будут рассмотрены более детально. ' Изостерическая регуляция активности ферментов осуществляется на уровне их каталитических центров. Реакционная способность и направленность работы каталитического центра прежде всего зависят от количества субстрата (закон действия масс). Интенсивность работы фермента определяется также наличием коферментов (для двухкомпонентных энзимов), кофакторов (специфически действующих катионов) и активаторов или ингибиторов, действующих на уровне каталитического центра. Активность тех или других ферментов может быть связана с конкуренцией за общие субстраты и коферменты, что является одним из способов взаимодействия различных метаболических циклов.

Некоторые ферменты, кроме каталитических (изостери-ческих) центров, имеют также аллостерические, т. е. расположенные в других местах рецепторные участки, которые служат для связывания аллостерических эффекторов (регуляторов). В качестве эффекторов могут выступать определенные метаболиты, гормоны или даже молекулы субстрата. В результате присоединения положительно или отрицательно действующего эффектора к аллостерическому центру происходит изменение всей структуры фермента (конформация), что приводит соответственно к активации или торможению функциональной активности каталитического центра.
Важным способом регуляции ферментативной активности является трансформация латентной формы фермента (зимогена) в активную форму. Это достигается разрушением некоторых ковалентных связей с помощью протеаз, восстановлением дисульфидных групп, фосфорилированием протеинкина-зами за счет АТР или ассоциацией неактивных субъединиц. Потенциально активные ферменты могут не функционировать из-за их компартментации, например, в лизосомах, причем освобождению лизосомных гидролаз способствуют кислые значения рН, свободнорадикальное окисление мембранных липидов и некоторые жирорастворимые витамины и стероиды. Инактивация ферментов осуществляется путем их связывания специфическими ингибиторами белковой природы, а также разрушения протеиназами.
Генетическая регуляция включает в себя регуляцию на уровне репликации, транскрипции, процессинга и трансляции. Молекулярные механизмы регуляции здесь те же (рН, ионы, модификация молекул, белки-регуляторы), однако сложность регуляторных сетей возрастает.
Роль генов состоит в хранении и передаче генетической информации. Информация записывается в структуре хромосомной ДНК в виде триплетного нуклеотидного кода. Информация в клетках передается благодаря синтезу РНК на матрице ДНК (транскрипция) и синтезу специализированных белков на матрице мРНК с участием рРНК и тРНК (трансляция). В ходе и после транскрипции или трансляции происходит модификация биополимеров (процес-синг), транспортирующихся в места назначения. Специализированные белковые молекулы в соответствии со своей «структурной» информацией путем самосборки образуют специфические комплексы, выполняющие различные функции: каталитические (ферменты), двигательные (сократительные белки), транспортные (насосы и переносчики), рецепторные (хемо-, фото- и механорецепторы), регуляторные (белковые активаторы, репрессоры, ингибиторы), защитные (лектины) и др.
Для извлечения в нужный момент необходимой информации из структур хромосом в клетке существует сложная система регуляции, не все стороны которой в настоящее время известны.
В качестве примера на рис. 2.2 приведена схема организации участка ядрышковой ДНК, кодирующей синтез трех рРНК: 18S, 5,8S и 25S (28S), 5S рРНК синтезируется в других участках хромосом. РНК-полимераза I, осуществляющая транскрипцию рибосомальных генов, присоединяется к про мотору (участку инициации транскрипции) после того, как все участки акцепторной зоны активируются соответствующими эффекторами (фитогормонами и др.). Транскрипция рибосо-мальных генов начинается с З'-конца ДНК. Структурные гены, кодирующие 18S, 5,8Sи 25S рРНК, разделены спейсерами (прокладками). В гене 25S рРНК наряду с кодирующими участками (экзонами) могут существовать некодирующие участки (интроны). Блоки структурных генов рРНК в ядрышковой ДНК, следуя друг за другом (тандемно), многократно повторяются.

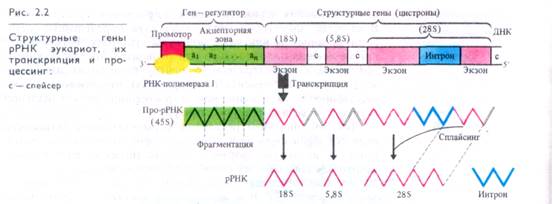
Зрелые рРНК образуются в ходе процессинга про-рРНК вследствие ее фрагментации и удаления участков, соответствующих спейсерам, а также благодаря сплайсингу, т. е. вырезанию интрона и объединению кодирующих участков рРНК. Интронная структура не характерна для рибосомальных генов, однако она представлена в генах хлоропластов, митохондрий, низших грибов и некоторых других эукариот.
Тандемная организация структурных генов у эукариот характерна для участков ДНК, кодирующих рРНК, тРНК и гистоны. Структурные гены мРНК, кодирующих синтез функционально взаимосвязанных ферментов, находятся в разных участках ДНК или даже в разных хромосомах.
Существует несколько гипотез, объясняющих механизмы постранскрипционной регуляции экспрессии генов мРНК. Согласно гипотезе Э. Дэвидсона и Р. Бриттена (1973, 1979), в ядерной ДНК наряду со структурными генами, кодирующими синтез различных мРНК, присутствуют интегрирующие регуляторные гены, содержащие многочисленные повторы (рис. 2.3). Если в нуклеоплазме в большом количестве появляются транскрипты с определенных регуля-торных генов (регуляторные транскрипты), то образуются комплементарные ассоциаты с соответствующими про-мРНК. В результате эти про-мРНК подвергаются быстрому про-цессингу, и активированные таким образом мРНК начинают функционировать. При этом резко возрастает скорость их считывания с соответствующих структурных генов.

На рис. 2.3 показано, каким образом эффекторы (сигналы), поступающие в ядро из цитоплазмы, индуцируют избирательную экспрессию генов.
Эффектор 1 (Э1, например фитогормон, взаимодействуя со своим рецепторным белком (РБ1), индуцирует транскрипцию определенного интегрирующего регуляторного гена с многочисленными повторами а. Про-РНК с этого регуляторного гена подвергается процессингу и регуляторные транскрипты, образуя комплементарные ассо-циаты с пром-РНКь индуцируют быстрое новообразование и активацию этой мРНК. В отличие от мРНКь считывание и активация мРНК2 (рис. 2.3) зависят от одновременного присутствия двух эффекторов. Причем один из них (Э1 может быть фитогормоном, а второй (Э2) — либо гормоном другого класса, либо метаболитом. Таким образом, синтез мРНК2 - пример генетической индукции, находящейся под двойным контролем. Так, синтез нитратредуктазы у растений можно индуцировать нитратом (субстратная индукция) и фитогормоном цитокинином, причем конечный продукт восстановления нитрата - аммоний - блокирует развитие нитрат-редуктазной активности.
Механизмы трансляции гораздо сложнее процессов транскрипции. В то время как транскрипцию обеспечивают десятки белков, для осуществления синтеза полинептида необходимы сотни специализированных белков. Только в рибосомах эукариот их 70—100, причем малая и большая рРНК выполняют роль каркаса, на котором осуществляется самосборка этих белков.
Интенсивность и направленность трансляции зависят от:
 1) концентрации информационных матриц, т. е. специфических мРНК, уровень которых определяется их синтезом, транспортом, хранением, активацией и распадом; 2) присутствия всех компонентов аппарата трансляции (рибосом, тРНК, аминокислот, АТР, GTP, синтетаз, регуляторных белков); 3) физико-химических условий (рН, ионов).
1) концентрации информационных матриц, т. е. специфических мРНК, уровень которых определяется их синтезом, транспортом, хранением, активацией и распадом; 2) присутствия всех компонентов аппарата трансляции (рибосом, тРНК, аминокислот, АТР, GTP, синтетаз, регуляторных белков); 3) физико-химических условий (рН, ионов).
Мембранная регуляция осуществляется благодаря сдвигам в мембранном транспорте, связыванию или освобождению ферментов и регуляторных белков и путем изменения активности мембранных ферментов. Все рассмотренные в гл. 1 функции мембран — транспортная, осмотическая, энергетическая и др. — одновременно являются и различными сторонами механизма регуляции внутриклеточного обмена веществ. Причем особое значение во всех этих механизмах имеет система мембранных хемо-, фото- и механорецепторов, позволяющих клетке оценивать качественные и количественные изменения во внешней и внутренней среде и в соответствии с этим изменять свойства мембран.
Контактная регуляция активности ферментов имеет место, в частности, в цистернах ЭР и в АГ, где идет достройка и модификация секретируемых белков. Связывание мембранами или освобождение ферментов, как уже отмечалось, также меняет их активность.
Дистанционная мембранная регуляция активности внутриклеточных ферментов осуществляется путем доставки субстратов и коферментов, удаления продуктов реакции, ионных и кисислотно-щелочных сдвигов в компартментах, фосфорилированием ферментов и другими способами. Для животных объектов существенную роль в регуляции активности некоторых ферментов играет аденилатциклазная система, локализованная в мембранах, и циклическая AMP. Однако присутствие этой системы в растительных клетках до настоящего времени остается дискуссионным. В то же время сдвиги в концентрации кальция выполняют в растительных клетках такую же регуляторную роль, как и в животных. Ионы Са2+, взаимодействуя с регуляторным белком кальмо-дулином, активируют протеинкиназы, фосфорилирующие различные белки, что приводит к изменению их функциональной активности. Са2+ специфически необходим для регуляции таких процессов, как движение цитоплазмы, митоз, секреция.
Мембранная регуляция генной активности на уровне репликации, транскрипции, процессинга и трансляции также осуществляется контактным и дистанционным способами. Причем с появлением у эукариот ядерной оболочки значение мембранной регуляции возросло. На возможность контактной регуляции указывают многочисленные связи хромосом эукариот с внутренней мембраной ядер, которые, по-видимому, принимают участие в пространственной организации хромосомного аппарата в нуклеоплазме. Примером контактной регуляции в цитоплазме может служить шероховатый рети-кулум, на поверхности мембран которого расположены рибосомы.
Дистанционная мембранная регуляция включает в себя транспорт предшественников синтеза ДНК, РНК и белков, освобождение регуляторных белков. Мембранное управление хромосомным и рибосомальным аппаратами осуществляется также с помощью ионных и кислотно-щелочных сдвигов. Так, активность РНК-полимеразы II, синтезирующей мРНК, повышается с увеличением ионной силы до 0,4 моль/л, причем необходимы Мп2+ и рН 7,5, в то время как ответственная за синтез рРНК РНК-полимераза I максимальную активность развивает при низкой ионной силе в присутствии Mg2+ и при рН 8,5. Эти данные указывают на то, что ионные отношения и рН, зависящие от регуляторной активности мембран, могут быть важным звеном в управлении генным аппаратом. Существует мнение, что сдвиги в ионном гомеостазе клеток в ответ на внешние воздействия служат первичным внутриклеточным индуктором процессов митоза и дифференциальной активности генов. В частности, сдвиг рН в кислую сторону в ряде случаев вызывает деление клеток. Синтез белков также зависит от качественного состава и количественного содержания ионов. Хорошо известна необходимость Mg2+ для сборки рибосом и полирибосом. Инициации трансляции благоприятствует низкая концентрация ионов К+ (NH4) порядка 30 — 50 ммоль/л и рН 7,4 — 7,6, а для последующего процесса элонгации полипептида в рибосомальном комплексе оптимальна повышенная концентрация К+ или NH4 (до 150 ммоль/л) и рН 8,3-8,4.
 Взаимодействие с помощью питательных веществ — наиболее простой способ связи между клетками, тканями и органами. У растений корни и другие гетеротрофные органы зависят от поступления ассимилятов, образующихся в листьях в процессе фотосинтеза.В свою очередь надземные части нуждаются в минеральных веществах и воде, поглощаемых корнями из почвы. Корни используют ассимиляты, поступающие из побега, на собственные нужды, а часть трансформированных органических веществ движется в обратном направлении. Изолированные корни в стерильных условиях для нормального развития помимо минеральных веществ и сахара нуждаются еще и в некоторых витаминах, таких, как В1 , В6 и никотиновая кислота. Очевидно, витамины поступают в корни из побегов.
Взаимодействие с помощью питательных веществ — наиболее простой способ связи между клетками, тканями и органами. У растений корни и другие гетеротрофные органы зависят от поступления ассимилятов, образующихся в листьях в процессе фотосинтеза.В свою очередь надземные части нуждаются в минеральных веществах и воде, поглощаемых корнями из почвы. Корни используют ассимиляты, поступающие из побега, на собственные нужды, а часть трансформированных органических веществ движется в обратном направлении. Изолированные корни в стерильных условиях для нормального развития помимо минеральных веществ и сахара нуждаются еще и в некоторых витаминах, таких, как В1 , В6 и никотиновая кислота. Очевидно, витамины поступают в корни из побегов.
Сдвиги в содержании различных элементов питания оказывают влияние на обмен веществ, физиологические и морфогенетические процессы у растений. Хорошо известны характерные изменения в растительном организме, обусловленные недостатком или избытком тех или иных минеральных веществ. При голодании усиливается конкуренция различных участков за продукты питания, что сказывается на процессах морфогенеза. При неблагоприятных условиях питания низшие организмы переходят к генеративным процессам. Цветение длиннодневных растений на длинном дне ускоряется при увеличении содержания углеводов и относительно низком содержании азотистых соединений, а зацветанию коротко-
дневных видов на коротком дне, наоборот, благоприятствует понижение содержания углеводов и увеличение азотистых соединений.
Однако нужно отметить, что трофическая регуляция носит скорее количественный, чем качественный, характер. При ограниченном питании у растений, как правило, развитие продолжается в соответствии с внутренними закономерностями, но у них формируются органы уменьшенного размера и сокращается количество листьев, плодов и семян. Интересно, что при этом конечная величина сформировавшихся семян, даже если это одно семя, мало отличается от нормы. Все это указывает, что наряду с трофическими взаимодействиями в растительном мире функционируют более совершенные системы регуляции, обеспечивающие взаимодействие всех его частей.
Фитогормоны — соединения, с помощью которых осуществляется взаимодействие клеток, тканей и органов и которые в малых количествах необходимы для запуска и регуляции физиологических и морфогенетических программ. Гормоны растений — сравнительно низкомолекулярные органические вещества (Мr 28 — 346). Они образуются в различных тканях и органах и действуют в очень низких концентрациях порядка 10-13-10-5 моль/л.
На возможность существования у растений веществ, функционально сходных с гормонами животных, впервые в 1880 г. указал Ч. Дарвин в своей книге «Способность к движению у растений». Исследуя двигательные реакции растений — тропизмы, он установил, что к воздействию внешних факторов наиболее чувствительны верхушки осевых органов растений (сенсорные зоны), а медленная двигательная ростовая реакция осуществляется в другом участке (в «моторной» зоне). В связи с этим он предположил, что раздражение (свет, гравитация и др.), воспринятое сенсорной зоной, передается в нижележащую моторную зону с помощью химическог стимула (рис. 2.4).

Г. Фиттинг, в 1909—1910 гг. изучавший особенности,опыления и оплодотворения у орхидей, обнаружил, что в поллиниях (комочки пыльцы орхидей) присутствует какое-то вещество, вызывающее разрастание завязи и формирование партенокарпических (бессемянных) плодов. Фиттинг первым предложил ввести в физиологию растений термин «гормон». Так как в дальнейшем исследования сосредоточились главным образом на изучении участия этих физиологически активных веществ в процессах роста и ростовых движений, их стали называть ростовыми гормонами или ростовыми веществами. За 60 лет с начала XX в. были открыты этилен, индолил-3-уксусная кислота (ауксин), цитокинины, гиббереллины, абсцизины, изучены фенольные ингибиторы роста негормонкльной природы. По мере открытия все новых фитогормонов и изучения разных аспектов их действия становилось очевидным, что физиологически активные вещества этого класса принимают участие не только в процессах роста, но и в созревании, старении, в явлениях стресса, транспорта веществ и во многих других.
Следует отметить, что гормональная система растений менее специализирована, чем у высокоорганизованных животных, имеющих обособленные эндокринные железы. В растительных организмах для включения и выключения морфогенетических и физиологических программ используются одни и те же фитогормоны в разных соотношениях.
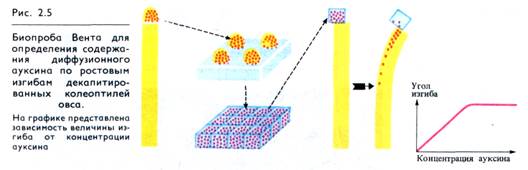
Предположение Дарвина о том, что верхушки колеоптилей проростков злаков синтезируют и выделяют вещество, способное активировать рост нижележащей зоны, было подтверждено в исследованиях первой четверти XX в. Ф. Вент выделил это ростовое вещество, накладывая отрезанные верхушки колеоптилей на агаровую пластинку. Ростовое вещество, диффундировавшее в агар, при наложении aгap-агаровых кубиков на срез декантированною колеоптиля вызывало ею pocт (рис. 2.5). В 1935 г. в лаборатории Ф. Кегля это вещество было идентифицировано как индолил-3-уксусная кислота (ИУК):

Соединения этой группы Кегль назвал ауксинами (от греч. «auxano» — расту). В последующие годы ИУК была обнаружена в различных растительных объектах.
 Метаболизм и транспорт. ИУК широко распространена в растительном мире от бактерий до высших растений. У высших растений ее больше всего в развивающихся почках и листьях, в активном камбии, в формирующихся семенах, в пыльце. В целом растении синтез ауксина обычно наиболее интенсивен в верхушке главного побега. Его предшественники — аминокислота триптофан или триптамин. ИУК может также освобождаться из связанного состояния (из гликозидов, комплексов с аминокислотами или белками).
Метаболизм и транспорт. ИУК широко распространена в растительном мире от бактерий до высших растений. У высших растений ее больше всего в развивающихся почках и листьях, в активном камбии, в формирующихся семенах, в пыльце. В целом растении синтез ауксина обычно наиболее интенсивен в верхушке главного побега. Его предшественники — аминокислота триптофан или триптамин. ИУК может также освобождаться из связанного состояния (из гликозидов, комплексов с аминокислотами или белками).
Транспорт ИУК в растительных тканях осуществляется полярно со скоростью 10—15 мм/ч от верхушки побега к корню (рис. 2.6). В стебле передвижение идет по живым клеткам проводящих пучков. Механизм полярного транспорта ауксина заключается в следующем: в апикальный конец клетки ИУК проникает пассивно вместе с ионами Н+ , а в базальном конце активно секретируется через клеточную мембрану. Активный транспорт ИУК специфически ингибируется N-1-нафтилфтала-миновой кислотой и морфактином.

Физиология и биохимия действия. Ауксин активирует деление и растяжение клеток, необходим для формирования проводящих пучков и корней, способствует разрастанию околоплодника. Ткани, обогащенные ауксином, обладают аттрагирующим действием, т. е. способны притягивать питательные вещества. В ряде случаев обработка ауксином задерживает процессы старения тканей и органов. Ауксин обусловливает явление апикального доминирования, т. е. тормозящее влияние апикальной почки на рост пазушных почек. Первостепенную роль играет ИУК в ростовых движениях — тропизмах и настиях (см. гл. 13).
Проникая в клетки, ИУК связывается со специфическими рецепторами, оказывая влияние на функциональную активность мембран, полирибосом и работу ядерного аппарата. Установлено, что в плазмалемме ауксин индуцирует работу Н+-помпы, в результате чего матрикс клеточных стенок закисляется. Это приводит к усилению активности кислых гидролаз и размягчению клеточных стенок, что является необходимым условием для роста клеток растяжением.
Включение Н+ - помпы усиливает поглотительную активность тканей, обогащенных ауксином (аттрагирующий эффект). Предполагается, что поступлеуше Са2+ в клетку способствует усилению секреции кислых гидролаз и полисахаридов, необходимых для дальнейшего роста клеточных стенок.
Комплекс фитогормона с рецептором, поступая в ядро, активирует синтез всех форм РНК, в том числе матричных, что в свою очередь приводит к формированию новых полирибосом и синтезу белков в цитоплазме (рис. 2.7). .
Вещества, необходимые для индукции деления растительных клеток, получили название цитюкининов. Впервые в чистом виде фактор клеточного деления был выделен из автоклави-рованного препарата ДНК спермы сельди (К. Миллер, Ф. Скуг и др., 1955). Это вещество было идентифицировано как 6-фурфуриламинопурин (кинетин). Природный цитокинин из незрелых зерновок кукурузы — зеатин — в 1963 г. получил Д. Летам:


Метаболизм и транспорт. В настоящее время цитокинины обнаружены у микроорганизмов, водорослей, папоротников, мхов и других высших растений. Наиболее богаты ими развивающиеся семена и плоды, а также меристематически активные участки. Установлено, что основное место синтеза цитокинина у вегетирующих растений — апикальные меристемы корней. Из корней цитокинины пассивно транспортируются в надземные органы по ксилеме (см. рис. 2.6).
Все природные цитокинины — производные изопентениладе-нина. Они синтезируются путем конденсации аденозин-5-моно-фосфага и Δ2-изопентенилпирофосфата. Цитокинин-активные рибонуклеозиды содержатся также в тРНК, однако эти тРНК сами по себе цитокининовой активностью не обладают. Связанные цитокинины в виде риботидов, рибозидов и гликозидов представляют собой транспортные и запасные формы.

Физиология и биохимия действия. Цитокинины индуцируют деление клеток, однако это их действие наблюдается лишь в присутствии ауксина. Обработка цитокинином вместе с ИУК побуждает дифференцированные клетки растений снова перейти к делению. У семядолей тыквы и листьев фасоли цитокинин активирует рост клеток растяжением. В культуре каллусной ткани цитокинин при повышенных концентрациях (0,5—1 мг/л) индуцирует образование побегов, у протонемы мха он способствует развитию гаметофора.
К. Мотес и сотрудники показали, что обогащение тканей цитокинином предотвращает распад хлорофилла и деградацию внутриклеточных структур у изолированных листьев. Эти же авторы установили, что участок изолированного листа, обработанный цитокинином (кинетином), становится зоной притяжения меченых аминокислот и других метаболитов (рис. 2.8). Таким образом, цитокинин необходим для нормального развития листа и для поддержания его аттрагирующей способности.

|
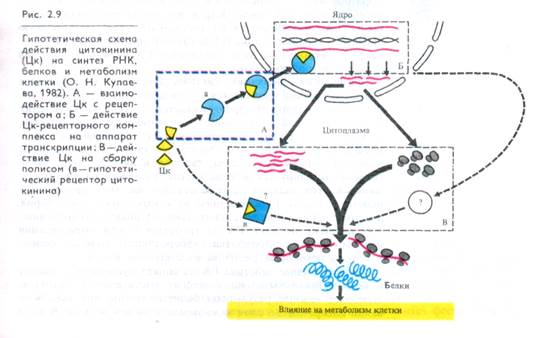
На молекулярном уровне цитокинин в комплексе со специфическим белковым рецептором усиливает активность РНК-полимеразы и матричную активность хроматина (рис. 2.9); при этом увеличиваются количество полирибосом и синтез белков, в том числе некоторых ферментов, в частности нитратредуктаны. Имеются данные о действии цитокинина на транспорт К+ , Н+ и Ca2+.
Японский исследователь Е. Куросава в 1926 г. установил, что культуральная жидкость фитопатогенного гриба Gibberella fujikuroi содержит химическое вещество, способствующее сильному вытягиванию стеблей у растений. Т. Ябута (1938) выделил это ещество в кристаллическом виде и назвал его гиббереллином. В 1954 г. англичанин Б. Кросс расшифровал структуру гибберелловой кислоты — тетрациклического дитерпеноида:

В настоящее время обнаружено более 60 различных гиббереллинов кислой и нейтральной природы. В связи с большим количеством открытых гиббереллинов был введен шифр ГА или просто А. Согласно этой номенклатуре гибберелловая кислота стала обозначаться как ГА3.
Метаболизм и транспорт. ГА и ГА-подобные вещества обнаружены у грибов, водорослей и высших растений. Наибольшее количество гиббереллинов у высших растений содержится в незрелых семенах. Гиббереллины синтезируются главным образом в листьях, а также в корнях. Свет стимулирует образование ГА. Транспорт ГА происходит пассивно с ксилем-ным и флоэмным током. Как и все полиизопреновые соединения, ГА синтезируются из ацетил-СоА через мевалоновую кислоту и геранилгераниол; ближайший предшественник ГА — каурен. Связанные в виде гликозидов ГА являются запасной и транспортной формами.
Физиология и биохимия действия. Действие ГА на растения наиболее типично проявляется в удлинении их стебля (рис. 2.10). Особенно наглядно это прослеживается у карликовых растений и у длиннодневных растений, находящихся в фазе розетки. У последних гиббереллин способствует образованию цветоноса, а часто и зацветанию. Места действия ГА — апикальные и интеркалярные меристемы, где под влиянием этого фито-гормона активируется деление клеток. Ускоряется также растяжение клеток, однако непосредственно на этот процесс ГА действует слабо. Гиббереллины не стимулируют рост корня, а в повышенных концентрациях даже ухудшают его состояние. Как правило, добавки ГА не требуется и при выращивании культуры тканей. Обработка гиббереллином выводит семена и клубни некоторых растений из состояния покоя.
Активирующее действие ГА на синтез нуклеиновых кислот и белков было выявлено у многих растительных объектов. Наиболее важные результаты были получены при использовании алейронового слоя эндосперма зерновок ячменя. В этом объекте под действием гиббереллина индуцируется синтез матричных РНК, которые кодируют образование ос-амилазы и некоторых других гидролаз. Как и другие фитогормоны, ГА прежде всего взаимодействует с цитоплазматическим рецептором белковой природы. Показано также, что гиббереллин активирует ферменты, ответственные за синтез фосфолипидов, входящих в состав мембран, образование гранулярного ЭР и секрецию ферментов. В тканях, обработанных гиббереллином, как правило, возрастает содержание ИУК.
В 1961 г. В. Лью и X. Карнс сухих зрелых коробочек хлопчатника выделили в кристаллическом виде вещество, ускоряющее опадение листьев, и назвали его абсцизином (от англ, abscission — отделение, опадение). Аналогичная работа была проделана в лаборатории Ф. Эддикотта; полученное ими вещество ускоряло опадение коробочек хлопчатника и ингиби-ровало рост отрезков колеоптилей, индуцированный ИУК. Ф. Уоринг и др. (1963) из листьев березы выделили ингибитор роста, который вызывал состояние покоя у почек растущих побегов, назвав это соединение дормином (от англ, dormancy — покой).


|