ИСТОРИЯ ЗЕМЛИ И ЖИЗНИ НА НЕЙ 10 страница
С другой стороны, несомненные пресноводные фауны (которые содержат, помимо всего прочего, дорожки следов на поверхности осадка) появляются намного позже - в девоне. В их состав входят скорпионы, мелкие (примерно в ладонь величиной) ракоскорпионы, рыбы и первые неморские моллюски; среди моллюсков есть и двустворчатые - длительно живущие организмы, неспособные переносить заморы и пересыхание водоемов. Фауны же с такими бесспорно-почвенными животными, как тригонотарбы ("панцирные пауки") и растительноядные двупарноногие многоножки существуют уже в силуре (лудловский век). А поскольку водная фауна всегда попадает в захоронения на порядок лучше, чем наземная, то все это позволяет сделать еще один вывод:
- Почвенная фауна появилась существенно раньше пресноводной. То есть - по крайней мере для животных пресные воды не могли играть роль "плацдарма" при завоевании суши.
Этот вывод, однако, заставляет нас вернуться к тому самому вопросу, с которого мы начинали свои рассуждения, а именно: вышли живые организмы на сушу или фактически создали ее как таковую? А.Г. Пономаренко (1993) полагает, что все сообщества, о которых шла речь выше, на самом деле трудно с определенностью назвать "наземными" или "сообществами внутриконтинентальных водоемов" (хотя по крайней мере маты должны были существенную часть времени находиться в воде). Он считает, что "существование настоящих континентальных водоемов, как текучих, так и стоячих, представляется весьма проблематичным до того, как в девоне сосудистая растительность несколько снизила скорость эрозии и стабилизировала береговую линию". Основные события должны были происходить в уже знакомых нам выположенных прибрежных амфибиотических ландшафтах без стабильной береговой линии - "не суша, не море" (см. главу 5 ).
Не менее необычная (с точки зрения сегодняшнего дня) обстановка должна была складываться и на водоразделах, занятых "первичными пустынями". В наши дни пустыни существуют в условиях недостатка влаги (при превышении испарения над выпадением осадков) - что и препятствует развитию растительности. А вот в отсутствие растений ландшафт парадоксальным образом становился тем более пустынным (по облику), чем больше выпадало осадков: вода активно размывала горные склоны, прорезая глубокие каньоны, при выходе на равнину давала конгломераты, а дальше по равнине распространялись разрозненные по поверхности псефиты [33], которые называют равнинным пролювием; ныне такие отложения слагают лишь конусы выноса временных водотоков.
Эта картина позволяет по-новому взглянуть на одно странное обстоятельство. Почти все известные силурийско-девонские наземные флоры и фауны найдены в различных точках древнего Континента красного песчаника (Old Red Sandstone), названного так по характерным для него породам - красноцветам; все местонахождения при этом связаны с отложениями, считающимися дельтовыми. Иными словами, выходит, что весь этот континент (объединявший Европу и восток Северной Америки) представляет собой как бы одну сплошную гигантскую дельту. Резонный вопрос: а где же располагались соответствующие реки - ведь для них на континенте таких размеров просто нет площадей водосбора! Остается предположить что все эти "дельтовые" отложения, по всей видимости, возникали именно в результате процессов эрозии в описанных выше "мокрых пустынях".
Итак, жизнь на суше (которая, впрочем, не вполне еще суша) вроде бы существует с незапамятных времен, а в конце силура просто-напросто появляется еще одна группа растений - сосудистые (Tracheophyta)... Однако на самом деле появление сосудистых растений - одно из ключевых событий в истории биосферы, ибо по своей средообразующей роли эта группа живых организмов не имеет себе равных, по крайней мере, среди эукариот. Именно сосудистая растительность и внесла, как мы увидим далее, решающий вклад в становление наземных ландшафтов современного облика.
Общепринятой считается та точка зрения, что некие водоросли, жившие вблизи берега, сначала "высунули голову на воздух", затем заселили приливно-отливную зону, а потом, постепенно превращаясь в высшие растения, целиком вышли на берег. За этим последовало постепенное завоевание ими суши. Предками высших растений большинство ботаников считает одну из групп зеленых водорослей - харовых (Charophyta); они образуют ныне сплошные заросли на дне континентальных водоемов - как пресных, так и соленых, тогда как в море (да и то лишь в опресненных заливах) найдены всего несколько видов. Харовые обладают дифференцированным талломом и сложно устроенными органами размножения; с высшими растениями их сближают несколько уникальных анатомических и цитологических признаков - симметричные спермии, присутствие фрагмопласта (структуры, участвующей в построении клеточной стенки в процессе деления) и наличие одинакового набора фотосинтетических пигментов и запасных питательных веществ.
Однако против этой точки зрения выдвигалось серьезное - чисто палеонтологическое - возражение. Если процесс преобразования водорослей в высшие растения действительно происходил в прибрежных водах (где условия для попадания в палеонтологическую летопись наиболее благоприятны), то почему мы не видим никаких его промежуточных стадий? Тем более, что и сами харовые появляются в позднем силуре - одновременно с сосудистыми растениями, а особенности биологии этой группы не дают оснований оснований предполагать для нее длительный период "скрытого существования"... Поэтому появилась парадоксальная, на первый взгляд, гипотеза: а почему, собственно говоря, появление макроостатков высших растений в конце силура должно однозначно трактоваться как следы их выхода на сушу? Может быть, совсем наоборот - это следы переселения высших растений в воду? Во всяком случае, многие палеоботаники (С.В. Мейен, Г. Стеббинс, Г. Хилл) активно поддерживали гипотезу о происхождении высших растений не от водных макрофитов (типа харовых), а от наземных зеленых водорослей. Именно этим сухопутным (а потому не имеющим реальных шансов попасть в захоронения) "первичным высшим растениям" могли принадлежать загадочные споры с трехлучевой щелью, которые весьма многочисленны в раннем силуре и даже в позднем ордовике (начиная с карадокского века).
Впрочем, недавно выяснилось, что, судя по всему, правы сторонники обеих точек зрения - каждые по-своему. Дело в том, что некоторые из микроскопических наземных зеленых водорослей имеют тот же самый комплекс тонких цитологических признаков, что харовые и сосудистые (см. выше); эти микроводоросли теперь стали включать в состав Charophyta. Таким образом, возникает вполне логичная и непротиворечивая картина. Первоначально существовала - на суше - группа зеленых водорослей ("микроскопические харовые"), от которой в силуре произошли две близкородственные группы: "настоящие" харовые, заселившие континентальные водоемы, и высшие растения, начавшие осваивать сушу, и лишь спустя некоторое время (в полном соответствии со схемой Мейена) появившиеся в прибрежных местообитаниях.
Из курса ботаники вам должно быть известно, что высшие растения (Embryophyta) делятся на сосудистые (Tracheophyta) и мохообразные (Bryophyta) - мхи и печеночники. Многие ботаники (например, Дж. Ричардсон, 1992) считают, что именно печеночники (если исходить из их современных жизненных стратегий) - главные претенденты на роль "первопроходцев суши": они живут ныне на наземных водорослевых пленках, в мелководных эфемерных водоемах, в почве - совместно с синезелеными водорослями. Интересно, что азотфиксирующая сине-зеленая водоросль Nostoc способна жить внутри тканей некоторых печеночников и антоцеротовых, обеспечивая своих хозяев азотом; это наверняка было очень важно для первых обитателей примитивных почв, где этот элемент не мог не быть в жестком дефиците [34]. Упоминавшиеся выше споры из позднеордовикских и раннесилурийских отложений наиболее схожи именно со спорами печеночников (достоверные макроостатки этих растений появляются позже, в раннем девоне).
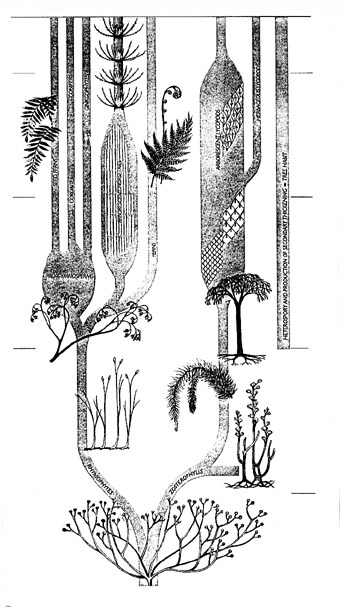
|
| РИСУНОК 30. Родословное древо первых наземных растений. |
Однако в любом случае мохообразные (если даже они действительно появились еще в ордовике) облик континентальных ландшафтов вряд ли изменили. Первые же сосудистые растения - риниофиты [35] - появились в позднем силуре (лудловский век); вплоть до раннего девона (жединский век) они были представлены крайне однообразными остатками единственного рода Cooksonia, простейшего и архаичнейшего из сосудистых. А вот в отложениях следующего века девона (зигенского) мы находим уже множество разнообразных риниофитов (рисунок 30). С этого времени среди них обособляются две эволюционные линии. Одна из них пойдет от рода Zosterophylum к плауновидным (в их число входят и древовидные лепидодендроны - одни из основных углеобразователей в следующем, карбоновом, периоде). Вторая линия (в ее основание обычно помещают род Psilophyton) ведет к хвощеобразным, папоротникам и семенным - голосеменным и покрытосеменным (рисунок 30). Даже девонские риниофиты еще очень примитивны и, честно говоря, неясно - можно ли назвать их "высшими растениями" в строгом смысле: у них есть проводящий пучок (правда, сложенный не трахеидами, а особыми вытянутыми клетками со своеобразным рельефом стенок), но отсутствуют устьица. Такое сочетание признаков должно свидетельствовать о том, что растения эти никогда не сталкивались с дефицитом воды (можно сказать, что вся их поверхность являет собою одно большое открытое устьице), и, по всей видимости, являлись гелофитами (то есть росли "по колено в воде", вроде нынешнего камыша).
Появление сосудистых растений с их жесткими вертикальными осями вызвало целый каскад экосистемных новаций, изменивших облик всей биосферы:
- Фотосинтезирующие структуры стали располагаться в трехмерном пространстве, а не на плоскости (как это было до сих пор - в период господства водорослевых корок и лишайников). Это резко увеличило интенсивность образования органического вещества и, тем самым, суммарную продуктивность биосферы.
- Вертикальное расположение стволов сделало растения более устойчивыми к занесению смываемым мелкоземом (по сравнению, например, с водорослевыми корками). Это уменьшило безвозвратные потери экосистемой неокисленного углерода (в виде органики) - совершенствование углеродного цикла.
- Вертикальные стволы наземных растений должны быть достаточно жесткими (по сравнению с водными макрофитами). Для обеспечения этой жесткости возникла новая ткань - древесина, которая после гибели растения разлагается относительно медленно. Таким образом, углеродный цикл экосистемы обретает дополнительное резервное депо и, соответственно, стабилизируется.
- Появление постоянно существующего запаса трудноразложимой органики (сконцентрированного в основном в почве) ведет к радикальной перестройке пищевых цепей. С этого времени большая часть вещества и энергии оборачивается через детритные, а не через пастбищные цепи (как это было в водных экосистемах).
- Для разложения трудноусваиваемых веществ, из которых состоит древесина - целлюлозы и лигнина - потребовались новые типы разрушителей мертвой органики. С этого времени на суше роль основных деструкторов переходит от бактерий к грибам.
- Для поддержания ствола в вертикальном положении (в условиях действия силы тяжести и ветров) возникла развитая корневая система: ризоидами - как у водорослей и мохообразных - тут уже не обойдешься. Это привело к заметному снижению эрозии и появлению закрепленных (ризосферных) почв.
С.В. Мейен полагает, что суша должна была покрыться растительностью к концу девона (зигенский век) [36], поскольку с начала следующего, карбонового, периода на Земле образуются уже практически все типы осадков, отлагающихся ныне на континентах. В дозигенские же времена континентальные осадки практически отсутствуют - видимо, в связи с их постоянным вторичным размывом в результате нерегулируемого стока. В самом начале карбона на континентах начинается угленакопление - а это свидетельствует о том, что на пути стока вод стояли мощные растительные фильтры. Не будь их, остатки растений непрерывно смешивались бы с песком и глиной, так что получались бы обломочные породы, обогащенные растительными остатками - углистые сланцы и углистые песчаники, а не настоящие угли.
Итак, возникшая в прибрежных амфибиотических ландшафтах густая "щетка" из гелофитов (можно назвать ее "риниофитный камыш") начинает действовать как фильтр, регулирующий плащевой сток: она интенсивно отцеживает (и осаживает) сносимый с суши обломочный материал и формирует тем самым стабильную береговую линию. Некоторым аналогом этого процесса может служить формирование крокодилами "аллигаторовых прудов": животные постоянно углубляют и расширяют населяемые ими болотные водоемы, выбрасывая грунт на берег. В результате их многолетней "ирригационной деятельности" болото превращается в систему чистых глубоких прудов, разделенных широкими облесенными "дамбами". Так и сосудистая растительность в девоне разделила пресловутые амфибиотические ландшафты на "настоящую сушу" и "настоящие пресноводные водоемы". Не будет ошибкой сказать, что именно сосудистая растительность стала истинным исполнителем заклинания: "Да будет твердь!" - отделивши оную твердь от хляби...
Именно с вновь возникшими пресноводными водоемами связано и появление в позднем девоне (фаменский век) первых тетрапод (четвероногих) - группы позвоночных, имеющих две пары конечностей; она объединяет в своем составе амфибий, рептилий, млекопитающих и птиц (попросту говоря, тетраподы - это все позвоночные, кроме рыб и рыбообразных). В настоящее время общепринято, что тетраподы ведут свое происхождение от кистеперых рыб (Rhipidistia) (рисунок 31); эта реликтовая группа имеет ныне единственного живого представителя, латимерию [37]. Достаточно популярная некогда гипотеза происхождения четвероногих от другой реликтовой группы рыб - двоякодышащих (Dipnoi), ныне практически не имеет сторонников.
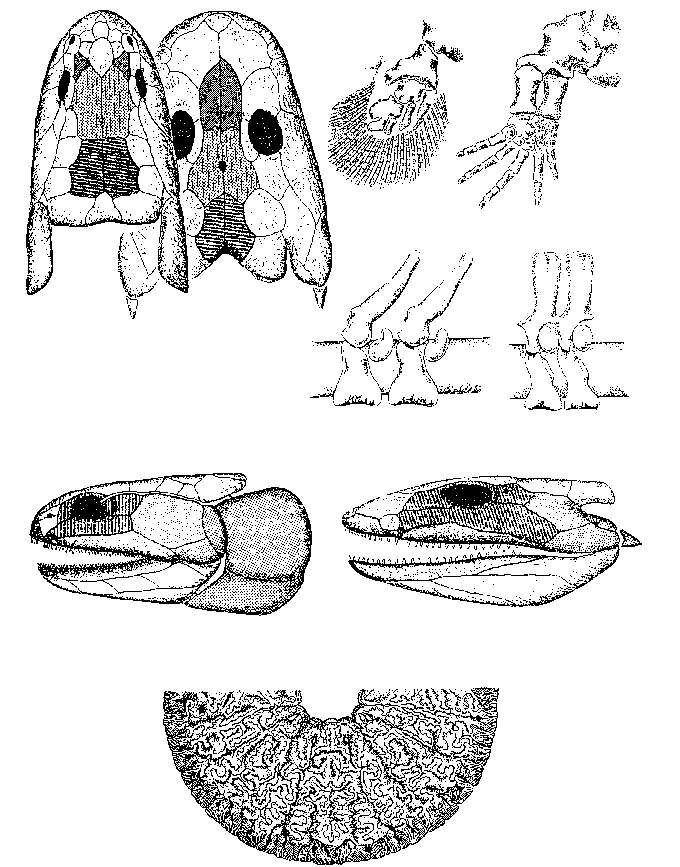
РИСУНОК 31. Преемственность анатомических структур (череп, конечности и позвонки) кистеперых рыб и амфибий: идентичные кости черепа имеют одинаковую заливку; идентичные кости обозначены одинаковыми буквами; отдельно - поперечный шлиф половинки зуба лабиритнтодонта.
Необходимо отметить, что в прежние годы возникновение ключевого признака тетрапод - двух пар пятипалых конечностей - считали их однозначной адаптацией к наземному (или по меньшей мере амфибиотическому) образу жизни. Ныне, однако, большинство исследователей склоняется к тому, что "проблема появления четвероногих" и "проблема их выхода на сушу" - суть вещи разные и даже не связанные между собою прямой причинной связью. Предки тетрапод жили в мелководных, часто пересыхающих, обильно заросших растительностью водоемах непостоянной конфигурации. Судя по всему, конечности появились для того, чтобы передвигаться по дну водоемов (это особенно важно, когда водоем обмелел настолько, что у тебя уже спина начинает вытарчивать наружу) и продираться сквозь плотные заросли гелофитов; особенно же полезны конечности оказались для того, чтобы при пересыхании водоема переползти посуху в другой, соседний.
Первые, девонские, тетраподы - примитивные амфибии лабиринтодонты [38] (название это происходит от их зубов с лабиринтоподобными складками эмали - структура, прямо унаследованная от кистеперых: см. рисунок 31), такие как ихтиостега и акантостега, в захоронениях всегда встречаются вместе с рыбами, которыми, судя по всему, и питались. Они были покрыты чешуей, как рыбы, имели хвостовой плавник (похожий на тот, что мы видим у сома или налима), органы боковой линии и - в некоторых случаях - развитый жаберный аппарат; конечность их еще не пятипалая (число пальцев достигает 8), и по типу сочленения с осевым скелетом - типично плавательная, а не опорная. Все это не оставляет сомнения в том, что существа эти были чисто водными (рисунок 32); если они и появлялись на суше при неких "пожарных" обстоятельствах (пересыхание водоема), то компонентом наземных экосистем наверняка не являлись [39]. Лишь много позже, в карбоновом периоде, появились мелкие наземные амфибии - антракозавры, которые, судя по всему, питались членистоногими, но об этом речь впереди (см. главу 10).

РИСУНОК 32. Архаичный лабиринтодонт Acanthostega : скелет (а) и реконструкция в естественной обстановке (б).
Особого внимания заслуживает тот факт, что в девоне появляется целый ряд неродственных параллельных групп стегоцефалоподобных кистеперых рыб - причем как до, так и после возникновения "настоящих" тетрапод (лабиринтодонтов). Одной из таких групп были пандерихтиды - кистеперые, лишенные спинного и анального плавников, чего не бывает ни у каких других рыб. По строению черепа (уже не "рыбьему", а "крокодильему"), плечевого пояса, гистологии зубов и положению хоан (внутренних ноздрей) пандерихтиды очень сходны с ихтиостегой, однако приобрели эти признаки явно независимо. Таким образом, перед нами процесс, который можно назвать параллельной тетраподизацией кистеперых (он был детально изучен Э.И. Воробьевой). Как обычно, "заказ" на создание четвероногого позвоночного, способного жить (или, по крайней мере, выживать) на суше был дан биосферой не одному, а нескольким "конструкторским бюро"; "выиграла конкурс" в конечном итоге та группа кистеперых, которая "создала" известных нам тетрапод современного типа. Однако наряду с "настоящими" тетраподами еще долго существовал целый спектр экологически сходных полуводных животных (типа пандерихтид), сочетающих признаки рыб и амфибий - если так можно выразиться, "отходы" процесса тетраподизации кистеперых.
9. Поздний палеозой - ранний мезозой: криоэры и термоэры. Палеозойские леса и континентальные водоемы - растения и насекомые.
Выдающийся палеоботаник С.В. Мейен, анализируя механизм возникновения прочно устоявшихся научных предрассудков, приводит в качестве примера цитату из школьного учебника биологии, где речь идет о карбоновом (каменноугольном) периоде "с его теплым влажным климатом и воздухом, богатым углекислым газом вследствие сильной вулканической деятельности... С конца каменноугольного периода в связи с усиленным горообразованием, охватившим в следующем (пермском) периоде весь земной шар, влажный климат почти повсеместно сменился сухим. В новых условиях древовидные папоротники стали быстро вымирать... Вымерли и семенные папоротники." Дело даже не в прямых фактических ошибках (например, "семенные папоротники" [40], которые якобы погибли из-за установления в перми засушливого климата, на самом деле благополучно дожили почти до конца мезозоя, а пермь вообще была временем их расцвета). Серьезнее другое: представления о том, что карбон - время теплого и влажного климата, а пермь - время повсеместной аридизации, сформировались в прошлом веке; эта картина была нарисована европейскими геологами и палеонтологами на европейском материале, а затем - безо всяких на то оснований! - распространена на всю Землю. Уже в начале нашего века стало ясно, что ситуация в Еврамерийской области карбона (территория нынешних Европы и Северной Америки) сходна с Катазией (Китай и Индокитай), но радикально отлична от того, что наблюдалось в Гондване (на материках Южного полушария и Индии) и в Ангариде (северная Азия) (см. рисунок 33); тем не менее, на возникший чуть ли не полтора века назад стереотип все это ничуть не повлияло - как можно видеть из цитированного выше учебника 80-х годов.
РИСУНОК 33. Расположение главных палеофлористических областей в позднем палеозое при современном положении материков (а) и на мобилистской реконструкции (б). А - Ангарская, Е - Еврвмерийская, К - Катазиатская и Г - Гондванская области.
Причины климатических различий, существовавших в позднем палеозое между Еврамерией и Катазией, с одной стороны, и Гондваной и Ангаридой - с другой, кажутся вполне очевидными: первые располагались на тогдашнем экваторе, а вторые - вблизи полюсов (рисунок 33, б). Представьте-ка себе, что мы попытаемся экстраполировать на всю нынешнюю Землю картину амазонских джунглей!.. Однако здесь сразу же возникает встречный вопрос: а всегда ли на Земле существовала широтная климатическая зональность, сходная с нынешней? Для ответа логично обратиться к сопоставлению высоко- и низкоширотных флор соответствующих эпох прошлого (учитывая при этом иное, чем теперь, расположение материков относительно полюса).
Картина эволюции растительности от девона до наших дней выяснена (в общих чертах) достаточно давно. Для наиболее молодых, кайнозойских, флор установлена ясная климатическая зональность, хотя и отличная от нынешней (на арктических островах росли деревья, характерные ныне для зоны широколиственных лесов - например, каштаны и платаны). Мезозойские флоры существенно более однообразны по всей Земле. Сложнее ситуация с палеозоем. Пермские и позднекарбоновые флоры Европы и Северной Америки, как уже было сказано, сходны с китайскими, но резко отличны и от сибирских, и от флор всех материков Южного полушария. Но ниже по геологическому разрезу - в раннем карбоне и далее, в девоне, мы снова сталкиваемся с единством флор различных материков. Отсюда можно заключить, что климатическая зональность была минимальной в девоне и начале карбона, затем усилилась в позднем палеозое, снова ослабла в мезозое, а потом опять начала усиливаться, достигнув ныне своего максимума.
Мы с вами помним, что в верхнепалеозойских слоях всех гондванских материков найдены ледниковые отложения - тиллиты (которые послужили одним из отправных пунктов в построениях Вегенера); следы оледенений найдены и в одновозрастных отложениях Ангариды. А вот за изъятием этого отрезка времени (поздний карбон - ранняя пермь) и современности климат в высоких широтах был если и не жарким, то во всяком случае безморозным: в раннем карбоне и Европы, и Шпицбергена, и Сибири найдены толстые, явно многолетние стволы плауновидных с маноксилической древесиной [41], а в эоцене острова Элсмир (Канадский архипелаг) - крокодилы. Современная климатическая картина с крупными полярными шапками из снега и льда - скорее исключение, чем правило в геологической истории. Так что следует искать ответа не на вопрос, почему в раннем карбоне и в мезозое не было полярных шапок, а на вопрос, отчего они иногда образовывались (и меняли весь климат планеты). Периоды существования такого контрастного климата с холодными полюсами, как в позднем палеозое и позднем кайнозое, называют криоэрами ("криос" - по-гречески холод), а выровненного по всей Земле (как в мезозое) - соответственно, термоэрами. Общее количество тепла, получаемое Землей от Солнца, считается достаточно постоянным во все времена; здесь существует своя циклика (см. главу 14), но расстояние-то между ними неизменно. Следовательно, дело в основном в распределении этого тепла по поверхности планеты, прежде всего - в характере и интенсивности теплопереноса от экватора к полюсам.
Для начала, как водится, несколько общих замечаний. Поскольку планета шарообразна, солнечные лучи всегда будут, при прочих равных, нагревать ее экватор сильнее, чем полюса - экваториально-полярный температурный градиент; любой градиент стремится к выравниванию (просто по Второму закону термодинамики) - в нашем случае за счет постоянного теплообмена между низкими широтами и высокими. Теплообмен этот осуществляется посредством конвекции в обеих подвижных оболочках Земли - гидросфере и атмосфере.
Конвекция в гидросфере - это теплые морские течения, которые обогревают высокоширотные области точно так же, как водяное отопление - ваши квартиры. Движущей силой конвекционных токов, как мы помним из главы 2 (о мантийной конвекции), являются возникающие в среде архимедовы силы плавучести: когда часть вещества "тонет" или "всплывает", этот объем - в силу связности среды - замещается веществом, поступающим из другой ее точки. В нашем случае токи в Мировом океане могут возникать за счет того, что "тонет" либо холодная (четырехградусная) вода в высоких широтах (термическая циркуляция), либо избыточно осолоненная (в результате испарения) вода на экваторе (галинная циркуляция). При термической циркуляции вода движется от экватора по поверхности, а от полюсов - по дну (формируя при этом холодную насыщенную кислородом психросферу), а при галинной - наоборот (рисунок 34, б).
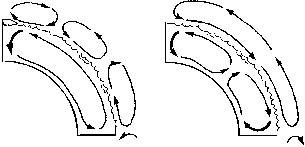
|
| РИСУНОК 34. Циркуляция в атмосфере и гидросфере: схема термической и галинной циркуляции в океанах: (а) - криоэра, (б) - термоэра. |
Говоря о конвекции в атмосфере, необходимо учитывать, что здесь тепло переносится главным образом водяным паром: тепловая энергия, затраченная на испарение воды, выделяется там, где этот пар, перенесенный воздушными течениями, превратится обратно в жидкость - то есть выпадет в виде осадков. Атмосфера каждого из полушарий распадается на три широтных сегмента [42] - конвективные ячейки: экваториальная, умеренных широт и приполярная. В каждой из ячеек существует относительно замкнутая воздушная циркуляция, причем направления циркуляции в граничащих между собой ячейках противоположны ("по часовой стрелке" - "против" - опять "по") - в точности, как в цепи шестеренок (рисунок 34, а). В одной половине ячейки доминируют восходящие токи, во второй - нисходящие; соответственно, влага, испаряющаяся в первой половине, выпадает главным образом во второй - и при этом происходит разгрузка теплоты парообразования. Например, в экваториальной ячейке Северного полушария ток направлен от севера к югу, так что в южной ее половине возникают влажные тропические леса, а в северной - засушливые саванны; в ячейке же умеренных широт, где направление тока обратное, пустыни возникают на юге, а субтропические и широколиственные леса - на севере. Другим фактором атмосферной конвекции (главным образом широтным) являются муссоны - сезонные ветры постоянного направления, дующие с океана на континент или обратно; с муссонами связано, среди прочего, чередование сухого сезона и сезона дождей в тропических широтах, где температура весь год постоянна.
Так вот, возвращаясь к крио- и термоэрам. Ныне (как, видимо, и вообще в криоэрах) основной приток тепла в высокие широты осуществляется мощными теплыми течениями вроде Гольфстрима. При этом возникает температурная аномалия: океан в районе 60-х широт существенно (почти на 20 градусов!) теплее, чем следовало бы из соображений геометрии планеты. Однако это обстоятельство имеет и оборотную сторону: на материке развивается мощный зимний антициклон (область высокого давления) с температурами на 20 градусов ниже, чем следовало бы. Из антициклона "вытекает" холодный сухой воздух, тогда как встречный ток теплого воздуха лишь "приподнимает верхушку" антициклона - то есть тепловая энергия расходуется на механическую работу против силы тяжести. В итоге континентальные антициклоны работают как мощные всепланетные холодильники (влияние Сибирского антициклона ощущается далеко на юге и приводит к холодным зимам не только в Приморье, но и в Китае, и даже во Вьетнаме), которые ослабляют муссоны и не дают им продвинуться сколь-нибудь далеко в полярном направлении. Теплые воды отводятся из эваториальной зоны столь быстро, что практически не успевают осолониться за счет испарения, так что галинная составляющая океанской циркуляции по сравнению с термической пренебрежимо мала; в атмосфере существуют упомянутые выше три ячейки. Ситуация может быть кратко охаректеризована так: "Водный теплоперенос - теплые океаны - холодные материки".
В термоэры (в частности, в мезозое) ситуация, судя по всему, была принципиально иной. А.Г. Пономаренко (1996), предложивший соответствующую модель, обращает внимание на два обстоятельства, кажущиеся необъяснимыми с нынешних позиций. Во-первых, приполярные области были очень теплыми (исходя из состава их фауны и флоры), что требует куда более интенсивного, чем ныне, теплопереноса от экватора к полюсу. Во-вторых, в тогдашней экваториальной зоне (где ныне находятся дождевые тропические леса) растительность была явно ксерофильной и возникали эвапориты - отложения, являющиеся показателем аридных (пустынных) условий.
При отсутствии полярных шапок температура океанической воды не падает до 4°, когда ее плотность максимальна, и не происходит полного опускания поверхностных вод. Компенсирующие такое погружение мощные теплые течения не возникают; теплая вода из экваториальной зоны практически не отводится, она осолоняется за счет испарения и погружается на дно - галинная циркуляция здесь резко доминирует над термической. Поверхностные слои океанов в умеренных широтах относительно холодны, и температурного контраста между океанами и материками не возникает; не возникает и зимнего антициклона, так что ничто теперь не мешает муссону переносить теплый воздух и водяной пар чуть ли не до самого полюса.
Если в криоэрах в каждом из полушарий существуют три атмосферные ячейки и единственная (термическая) океанская, то в термоэрах ситуация, похоже, была как бы зеркальной. В атмосфере существует единственная ячейка, в которой происходит прямой перенос тепла и влаги из приэкваториальных областей в приполярные; это хорошо объясняет упомянутый выше парадокс с располагающимся на экваторе аридным поясом [43]. В гидросфере, напротив, существуют две ячейки, причем в экваториальной ячейке циркуляция идет по галинному типу, а в высокоширотной - по термическому. В зоне соприкосновения гидросферных ячеек, где водные массы движутся друг навстречу другу, должны возникать апвелинги [44]; подтверждением тому служит распространение по этой предполагаемой границе обогащенных органикой черносланцевых формаций, которые отлагаются в избыточно-продуктивных морских акваториях.