Функции варолиева моста.
Варолиев мост выполняет двигательные, сенсорные, интегративные и проводниковые функции. Важные функции моста связаны с наличием в нем ядер черепных нервов.
V пара – тройничный нерв (смешанный). Двигательное ядро нерва иннервирует жевательные мышцы, мышцы нёбной занавески и мышцы, напрягающие барабанную перепонку. Чувствительное ядро получает афферентные аксоны от рецепторов кожи лица, слизистой оболочки носа, зубов, 2/3 языка, надкостницы костей черепа, конъюнктивы глазного яблока.
VI пара – отводящий нерв (двигательный), иннервирует прямую наружную мышцу, отводящую глазное яблоко кнаружи.
VII пара – лицевой нерв (смешанный), иннервирует мимические мышцы лица, подъязычную и подчелюстную слюнные железы, передает информацию от вкусовых рецепторов передней части языка.
VIII пара – преддверно-улитковый (чувствительный). Улитковая часть преддверно-улиткового нерва заканчивается в мозге в улитковых ядрах; преддверная его часть – в треугольном ядре, ядре Дейтерса, ядре Бехтерева. Здесь происходит первичный анализ вестибулярных раздражений их силы и направленности.
В мосте располагается пневмотаксический центр, запускающий центр выдоха продолговатого мозга, а также группа нейронов, активирующих центр вдоха.
Через мост проходят все восходящие и нисходящие пути, связывающие мост с мозжечком, спинным мозгом, корой больших полушарий и другими структурами центральной нервной системы. По мостомозжечковым проводящим путям через мост осуществляется контролирующее влияние коры полушарий большого мозга на мозжечок.
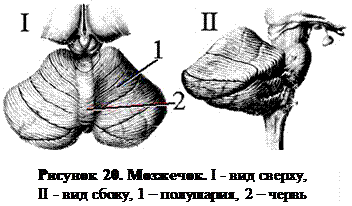 2. 3. 3 Мозжечок (cerebellum).Мозжечок (малый мозг) располагается сзади от моста и продолговатого мозга. Он состоит из средней, непарной, филогенетически старой части – червя – и парных полушарий, свойственных лишь млекопитающим. Полушария мозжечка развиваются параллельно с корой больших полушарий и достигают у человека значительных размеров. Червь с нижней стороны погружен глубоко между полушариями; верхняя его поверхность переходит в полушария постепенно (рис. 20).
2. 3. 3 Мозжечок (cerebellum).Мозжечок (малый мозг) располагается сзади от моста и продолговатого мозга. Он состоит из средней, непарной, филогенетически старой части – червя – и парных полушарий, свойственных лишь млекопитающим. Полушария мозжечка развиваются параллельно с корой больших полушарий и достигают у человека значительных размеров. Червь с нижней стороны погружен глубоко между полушариями; верхняя его поверхность переходит в полушария постепенно (рис. 20).
В целом мозжечок имеет обширные эфферентные связи со всеми двигательными системами стволовой части мозга: кортикоспинальной, руброспинальной, ретикулоспинальной и вестибулоспинальной. Не менее разнообразными являются и афферентные входы мозжечка.
Со стволовой частью мозга мозжечок соединяется тремя парами ножек. Самые толстые средние ножки расширяясь, переходят в варолиев мост. Верхние ножки начинаются в зубчатых ядрах мозжечка (см. ниже) и направляются к четверохолмию среднего мозга. Третья пара ножек (нижняя) спускается вниз, сливаясь с продолговатым мозгом. Афферентные волокна, приходящие в мозжечок, преимущественно входят в состав средних и нижних ножек, тогда как эфферентные собраны главным образом в верхних ножках мозжечка.
 Вся поверхность мозжечка разделяется глубокими бороздами на доли. В свою очередь, каждая доля параллельными бороздками разделяется на извилины; группы извилин формируют дольки мозжечка. Каждую дольку обозначают как классическим названием (язычок, центральная, вершина и т. д.), так и латинской нумерацией (I – Х) в соответствии с распространенной номенклатурой.
Вся поверхность мозжечка разделяется глубокими бороздами на доли. В свою очередь, каждая доля параллельными бороздками разделяется на извилины; группы извилин формируют дольки мозжечка. Каждую дольку обозначают как классическим названием (язычок, центральная, вершина и т. д.), так и латинской нумерацией (I – Х) в соответствии с распространенной номенклатурой.
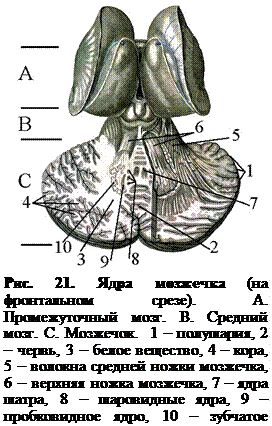
Полушария и червь мозжечка состоят из лежащего на периферии серого вещества – коры – и расположенного глубже белого вещества, в котором заложены скопления нервных клеток, образующие ядра мозжечка – ядра шатра, шаровидные, пробковидные и зубчатые (рис. 21).
 Кора мозжечка имеет специфическое, нигде в ЦНС не повторяющееся, строение. Она представлена тремя слоями (рис. 22). Самый поверхностный слой – молекулярный, он состоит из параллельных волокон и разветвлений дендритов и аксонов нейронов нижележащих слоев. В нижней части молекулярного слоя расположены тела корзинчатых клеток, аксоны которых оплетают тела и начальные сегменты аксонов клеток Пуркинье. Здесь же в молекулярном слое имеется некоторое количество звездчатых клеток.
Кора мозжечка имеет специфическое, нигде в ЦНС не повторяющееся, строение. Она представлена тремя слоями (рис. 22). Самый поверхностный слой – молекулярный, он состоит из параллельных волокон и разветвлений дендритов и аксонов нейронов нижележащих слоев. В нижней части молекулярного слоя расположены тела корзинчатых клеток, аксоны которых оплетают тела и начальные сегменты аксонов клеток Пуркинье. Здесь же в молекулярном слое имеется некоторое количество звездчатых клеток.
Во втором слое коры – ганглиозном – сосредоточены тела клеток Пуркинье. Эти крупные клетки ориентированы вертикально по отношению к поверхности коры мозжечка. Их дендриты поднимаются вверх и широко ветвятся в молекулярном слое. Дендриты клеток Пуркинье содержат множество шипиков, на которых образуют синапсы параллельные волокна молекулярного слоя. Аксоны клеток Пуркинье спускаются к ядрам мозжечка. Часть из них заканчивается на вестибулярных ядрах. Аксоны клеток Пуркинье представляют собой практически единственный выход из коры мозжечка.
Под ганглиозным слоем лежит гранулярный (зернистый) слой, который содержит большое число тел клеток-зерен, или гранулярных клеток. По некоторым подсчетам их число может достигать 10 млрд. Аксоны клеток-зерен поднимаются вертикально вверх в молекулярный слой и там Т-образно ветвятся. Ветви идут параллельно поверхности коры и образуют синапсы на дендритах других клеток. Здесь же в гранулярном слое лежат клетки Гольджи, аксоны которых подходят к клеткам-зернам.
Все клетки коры мозжечка являются тормозящими за исключением клеток-зерен, которые оказывают возбуждающее воздействие.
В мозжечок поступает два типа волокон. Это, во-первых, лазающие, или лиановидные, идущие из нижних олив продолговатого мозга. Нижняя олива получает афферентные импульсы от кожных рецепторов, мышц, суставных оболочек, надкостницы по так называемым спинномозжечковым трактам: заднему (дорсальному) и переднему (вентральному). Лазающие волокна широко ветвятся и подобно лианам оплетают дендриты клеток Пуркинье, формируя на них синапсы. Возбуждая клетки Пуркинье, они усиливают тормозное влияние этих клеток на ядра моста.
Вторая система афферентных волокон – это мшистые, или моховидные волокна, идущие от ядер моста (в которые также поступает информация от рецепторов мышц, сухожилий, вестибулярных ядер) и оканчивающиеся на клетках-зернах. Клетки-зерна посылают нервные импульсы на корзинчатые, звездчатые клетки, клетки Гольжи, возбуждая их, что приводит к торможению активности клеток Пуркинье (снятию тормозного влияния коры на ядра мозжечка).
Таким образом, путь, идущий на клетки Пуркинье через лиановидные волокна, пособствует усилению тормозного влияния клеток Пуркинье на ядра мозжечка, а путь, идущий через моховидные волокна, наоборот, снимает это тормозное влияние.
Следовательно, деятельность всей нейрональной системы коры мозжечка сводится к торможению ядер, над которыми кора надстроена. Согласно гипотезе, высказанной Дж. Экклсом, большое количество тормозных нейронов в коре мозжечка предотвращает длительную циркуляцию возбуждения по нейронным цепям. Любой возбуждающий импульс, приходя в кору мозжечка, превращается в торможение за время порядка 100 мс. Так происходит как бы автоматическое стирание предшествующей информации, которое позволяет коре мозжечка участвовать в регуляции быстрых движений.
С точки зрения функций, которые выполняет мозжечок, его делят на три части: архиоцеребеллум (древний мозжечок), палеоцеребеллум (старый мозжечок) и неоцеребеллум (новый мозжечок).
Кора архиоцеребеллума (внутренняя часть – червь мозжечка) связана с ядром шатра, которое регулирует активность вестибулярных ядер. Поэтому архиоцеребеллум является вестибулярным регулятором. Повреждения древних структур мозжечка приводят к нарушению равновесия.
Функция палеоцеребеллума, или средней части коры – взаимная координация позы и целенаправленного движения, а также коррекция выполнения сравнительно медленных движений по механизму обратной связи – реализуется с участием пробковидных и шаровидных ядер. При повреждении структур старого мозжечка больным трудно стоять и ходить, особенно в темноте, при отсутствии зрительной коррекции.
Неоцеребеллум (латеральная часть коры мозжечка) совместно с зубчатым ядром играет важную роль в программировании сложных движений, выполнение которых идет без использования механизма обратных связей. В итоге возникает целенаправленное движение, выполняемое с большой скоростью, например, игра на фортепиано. При нарушении структур неоцеребеллума нарушаются сложные последовательности движений, они становятся аритмичны и замедлены.
Функции мозжечка изучались в клинике при его поражениях у человека, а также у животных путем удаления (экстирпации мозжечка) (Л. Лючиани, Л. А. Орбели). В результате выпадения функций мозжечка возникают двигательные расстройства (на стороне поражения), которые итальянский физиолог Л. Лючиани охарактеризовал знаменитой триадой А – астазия, атония и астения. Последующие исследователи добавили еще один симптом – атаксия (тетрада А).
Астазия – утрата способности к длительному сокращению мышц, что затрудняет стояние, сидение и т. д., невозможность сохранения неподвижного положения, непрерывные качательные движения, дрожание головы, туловища и конечностей.
Атония – резкое падение и неправильное распределение тонуса мышц.
Астения – снижение силы мышечного сокращения, быстрая утомляемость мышц.
Атаксия – нарушение координации движений. Здесь ярче всего проявляется невозможность выполнения движений в нужном порядке, в определенной последовательности. Проявлениями атаксии являются также адиадохокинез, асинергия, пьяная шаткая походка. При адиадохокинезе человек не способен быстро вращать ладони вниз-вверх. При асинергии мышц он не способен сесть из положения лежа без помощи рук. Пьяная походка характеризуется тем, что человек ходит, широко расставив ноги, шатаясь из стороны в сторону от линии ходьбы. Врожденных двигательных актов у человека не так уж много (например, сосание), большинство же движений он выучивает в течение жизни, и они становятся автоматическими (ходьба, письмо и т. д.). Когда нарушается функция мозжечка, движения становятся неточными, негармоничными, разбросанными, часто не достигают цели.
Дисметрия – несоответствие между интенсивностью мышечного сокращения и задачей выполняемого движения, что может выражаться в понижении или повышении интенсивности рефлекторных реакций. Например, человек, поднимаясь по лестнице, слишком высоко поднимает колени («петушиная походка»), или наоборот, спотыкается о каждую ступеньку.
Дизартрия – расстройство организации речевой моторики. При повреждении мозжечка речь больного становится растянутой, слова иногда произносятся как бы толчками (скандированная речь).
С течением времени двигательные расстройства сглаживаются. Только предвзятое наблюдение выявляет некоторые нарушения (фаза компенсации). Как показал Э. А. Асратян, компенсация функций происходит за счет коры головного мозга. Мозжечок участвует в регуляции движений, делая их плавными, точными, соразмерными.
Мозжечок оказывает влияние также на ряд вегетативных функций, например желудочно-кишечного тракта, на уровень кровяного давления, на состав крови.
Долгое время мозжечок считался структурой, ответственной исключительно за координацию движений. Сегодня ученые все чаще говорят о его участии в процессах восприятия и когнитивной деятельности. Так, нейробиологи, изучающие когнитивные функции головного мозга, обнаружили, что у людей эта структура сохраняет высокую активность во время разнообразных форм деятельности, не связанных непосредственно с движениями.
 2. 3. 4 Средний мозг (mesencephalon ). Средний мозг расположен над мостом и представлен ножками мозга и четверохолмием. Ножки мозга состоят из основания и покрышки, между которыми находится черная субстанция, содержащая сильно пигментированные клетки. В покрышке мозга располагаются ядра блокового (IV пара) и глазодвигательного (III пара) нервов. Полость среднего мозга представлена узким каналом – сильвиевым водопроводом, который соединяет III и IV мозговые желудочки. Длина среднего мозга у взрослого человека около 2 см, вес – 26 г. В процессе эмбрионального развития средний мозг формируется из среднего мозгового пузыря, боковые выпячивания которого перемещаются латерально и образуют сетчатку глаза, которая структурно и функционально представляет собой вынесенный на периферию нервный центр среднего мозга.
2. 3. 4 Средний мозг (mesencephalon ). Средний мозг расположен над мостом и представлен ножками мозга и четверохолмием. Ножки мозга состоят из основания и покрышки, между которыми находится черная субстанция, содержащая сильно пигментированные клетки. В покрышке мозга располагаются ядра блокового (IV пара) и глазодвигательного (III пара) нервов. Полость среднего мозга представлена узким каналом – сильвиевым водопроводом, который соединяет III и IV мозговые желудочки. Длина среднего мозга у взрослого человека около 2 см, вес – 26 г. В процессе эмбрионального развития средний мозг формируется из среднего мозгового пузыря, боковые выпячивания которого перемещаются латерально и образуют сетчатку глаза, которая структурно и функционально представляет собой вынесенный на периферию нервный центр среднего мозга.
Наиболее крупными ядрами среднего мозга являются красные ядра, черная субстанция, ядра черепных (глазодвигательного и блокового) нервов и ядра ретикулярной формации. Через средний мозг проходят восходящие пути к таламусу, большим полушариям и мозжечку, и нисходящие пути к продолговатому и спинному мозгу.
В среднем мозге располагается большое количество нейронов ретикулярной формации. В четверохолмии выделяют верхнее и нижнее двухолмия (рис. 23).
Средний мозг выполняет ряд функций: проводниковую, двигательную и рефлекторную.
Проводниковая функция заключается в том, что через него проходят все восходящие пути к вышележащим отделам: таламусу (медиальная петля, спииноталамический путь), большому мозгу и мозжечку. Нисходящие пути идут через средний мозг к продолговатому и спинному мозгу. Это пирамидный путь, корково-мостовые волокна, руброретикулоспинальный путь.
Двигательная функцияреализуется за счет ядра блокового нерва (n. trochlearis), ядер глазодвигательного нерва (п. oculomotorius), красного ядра (nucleus ruber), черного вещества (substantia nigra).
Красные ядра располагаются в покрышке (рис. 23.– В, 4). Будучи составной частью организующей движения экстрапирамидной системы, красные ядра получают нервные импульсы от моторной коры, подкорковых ядер, ядер мозжечка, черной субстанции среднего мозга и дают начало руброспинальному (красноядерно-спинномозговому) тракту, который как общий путь обеспечивает регуляцию тонуса скелетных мышц, снижая тонус мышц-разгибателей. Об этом свидетельствует классический опыт Ч. Шеррингтона с перерезкой стволовой части мозга. Если перерезку произвести на уровне переднего края задних холмов четверохолмия и таким образом отделить красное ядро от заднего мозга, то у кошки развивается децеребрационная ригидность. Это состояние характеризуется сильным напряжением мышц-разгибателей конечностей, шеи, спины. Основной причиной возникновения децеребрационной ригидности служит выраженное активирующее влияние латерального вестибулярного ядра (ядро Дейтерса) на мотонейроны разгибателей. Это влияние максимально в отсутствие тормозных влияний красного ядра, вышележащих структур и мозжечка. При перерезке мозга ниже ядра латерального вестибулярного нерва децеребрационная ригидность исчезает.
Красные ядра, получая информацию от двигательной зоны коры большого мозга, подкорковых ядер и мозжечка о готовящемся движении и состоянии опорно-двигательного аппарата, посылают корригирующие импульсы к мотонейронам спинного мозга по руброспинальному тракту и тем самым регулируют тонус мускулатуры, подготавливая его уровень к намечающемуся произвольному движению.
Со средним мозгом связано осуществление выпрямительных и статокинетических рефлексов. Выпрямительные рефлексы состоят из двух фаз: подъема головы и последующего подъема туловища. Первая фаза осуществляется вследствие рефлекторных влияний с рецепторов вестибулярного аппарата и кожи, вторая – с проприорецепторов мышц шеи и туловища. Статокинетические рефлексы направлены на возвращение тела в исходное положение при перемещении тела в пространстве, при вращении.
Другое функционально важное ядро среднего мозга – черная субстанция (Земмеринга) (рис. 23. – 5). Оно связано с лежащими в основании полушарий переднего мозга базальными ганглиями – полосатым телом и бледным шаром – и регулирует акты жевания, глотания (их последовательность), обеспечивает точные движения пальцев кисти руки, например при письме. Нейроны этого ядра способны синтезировать медиатор дофамин, который поставляется аксональным транспортом к базальным ганглиям головного мозга. Поражение черного вещества приводит к нарушению пластического тонуса мышц. Тонкая регуляция пластического тонуса при игре на скрипке, письме, выполнении графических работ обеспечивается черным веществом. В то же время при длительном удержании определенной позы происходят пластические изменения в мышцах за счет изменения их коллоидных свойств, что обеспечивает наименьшие затраты энергии. Регуляция этого процесса осуществляется клетками черного вещества.
Повреждение черной субстанции, вызывающее дегенерацию дофаминэргических путей к полосатому телу, связано с тяжелым неврологическим заболеванием – болезнью Паркинсона. Паркинсонизм проявляется в нарушении тонких содружественных движений, функции мимической мускулатуры и в появлении непроизвольных мышечных сокращений, или тремора. Этот болезненный синдром может быть снят при введении L-диоксифенилаланина – вещества, из которого синтезируется дофамин в организме.
Таким образом, восполняя дефицит медиатора, стало возможным купировать неврологическое заболевание и вместе с тем привести фактические доказательства роли черной субстанции среднего мозга в сенсомоторной координации движений.
Нейроны ядер глазодвигательного и блокового нервов регулируют движение глаза вверх, вниз, наружу, к носу и вниз к углу носа. Нейроны добавочного ядра глазодвигательного нерва (ядро Якубовича) регулируют просвет зрачка и кривизну хрусталика.
Рефлекторные функции. Функционально самостоятельными структурами среднего мозга являются бугры четверохолмия. Верхние из них являются первичными подкорковыми центрами зрительного анализатора (вместе с латеральными коленчатыми телами промежуточного мозга), нижние – слухового (вместе с медиальными коленчатыми телами промежуточного мозга). В них происходит первичное переключение зрительной и слуховой информации. От бугров четверохолмия аксоны их нейронов идут к ретикулярной формации ствола, мотонейронам спинного мозга. Нейроны четверохолмия могут быть полимодальными и детекторными. В последнем случае они реагируют только на один признак раздражения, например смену света и темноты, направление движения светового источника и т. д. Основная функция бугров четверохолмия – организация реакции настораживания и так называемых старт-рефлексов на внезапные, еще не распознанные, зрительные (верхнее двухолмие) или звуковые (нижнее двухолмие) сигналы. Активация среднего мозга в этих случаях через гипоталамус приводит к повышению тонуса мышц, учащению сокращений сердца; происходит подготовка к избеганию или к оборонительной реакции.
Четверохолмие организует ориентировочные зрительные и слуховые рефлексы.
У человека четверохолмный рефлекс является сторожевым. В случаях повышенной возбудимости четверохолмий при внезапном звуковом или световом раздражении у человека возникает вздрагивание, иногда вскакивание на ноги, вскрикивание, максимально быстрое удаление от раздражителя, подчас безудержное бегство. При нарушении четверохолмного рефлекса человек не может быстро переключаться с одного вида движения на другое. Следовательно, четверохолмия принимают участие в организации произвольных движений.
2. 3. 5 Промежуточный мозг (diencephalon).Промежуточный мозг располагается под мозолистым телом и сводом, срастаясь по бокам с полушариями большого мозга. К нему относятся: таламус (зрительные бугры), гипоталамус (подбугорная область), эпиталамус (надбугорная область) и метаталамус (забугорная область). Полостью промежуточного мозга является III желудочек мозга.
Эпиталамусвключает в себя железу внутренней секреции – эпифиз (шишковидное тело). В темноте она вырабатывает гормон мелатонин, который тормозит половое созревание, а также влияет на рост скелета.
Метаталамус представлен латеральными и медиальными коленчатыми телами. Латеральное, или наружное, коленчатое тело – это подкорковый центр зрения имеет прямые эфферентные связи с затылочной долей коры большого мозга и афферентные связи с сетчаткой глаза и с передними буграми четверохолмий. Нейроны латеральных коленчатых тел по-разному реагируют на цветовые раздражения, включение, выключение света, т. е. могут выполнять детекторную функцию.
 Медиальное коленчатое тело – подкорковый, таламический центр слуха, в него поступают афферентные импульсы из латеральной петли и от нижних бугров четверохолмий. Эфферентные пути от медиальных коленчатых тел идут в височную долю коры большого мозга, достигая там первичной слуховой зоны. Медиальное коленчатое тело имеет четкую тонотопичность. Следовательно, уже на уровне промежуточного мозга обеспечивается пространственное распределение чувствительности всех сенсорных систем организма, в том числе сенсорных посылок от интерорецепторов сосудов, органов брюшной, грудной полостей.
Медиальное коленчатое тело – подкорковый, таламический центр слуха, в него поступают афферентные импульсы из латеральной петли и от нижних бугров четверохолмий. Эфферентные пути от медиальных коленчатых тел идут в височную долю коры большого мозга, достигая там первичной слуховой зоны. Медиальное коленчатое тело имеет четкую тонотопичность. Следовательно, уже на уровне промежуточного мозга обеспечивается пространственное распределение чувствительности всех сенсорных систем организма, в том числе сенсорных посылок от интерорецепторов сосудов, органов брюшной, грудной полостей.
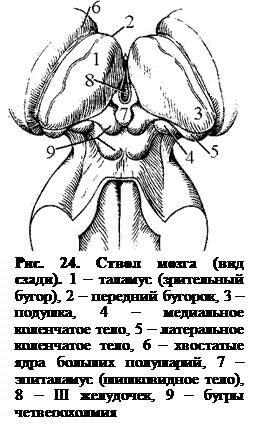
Таламус(thalamus, зрительный бугор) – парный орган яйцевидной формы (рис. 24) передняя часть которого заострена (передний бугорок), а задняя расширенная часть (подушка) нависает над коленчатыми телами. Медиальная поверхность таламуса обращена в полость III желудочка мозга.
Таламус – чувствительное ядро подкорки. Его называют «коллектором чувствительности», так как к нему сходятся афферентные (чувствительные) пути от всех рецепторов, исключая обонятельные.
В ядрах таламуса происходит переключение информации, поступающей от экстеро-, проприо- и интероцепторов на начинающиеся здесь таламокортикальные пути.
Главной функцией таламуса является интеграция (объединение) всех видов чувствительности. Для анализа внешней среды недостаточно сигналов от отдельных рецепторов. В таламусе происходит сопоставление информации, получаемой по различным каналам, и оценка ее биологического значения. В зрительном бугре насчитывается около 40 пар ядер, которые подразделяются на специфические (на нейронах этих ядер заканчиваются восходящие афферентные пути), неспецифические (ядра ретикулярной формации) и ассоциативные. Через ассоциативные ядра таламус связан со всеми двигательными ядрами подкорки – полосатым телом, бледным шаром, гипоталамусом и с ядрами среднего и продолговатого мозга.
От специфических ядер информация о характере сенсорных стимулов поступает в строго определенные участки III – IV слоев коры большого мозга (соматотопическая локализация). Нарушение функции специфических ядер приводит к выпадению конкретных видов чувствительности, так как ядра таламуса, как и кора большого мозга, имеют соматотопическую локализацию. Отдельные нейроны специфических ядер таламуса возбуждаются рецепторами только своего типа. К специфическим ядрам таламуса идут сигналы от рецепторов кожи, глаз, уха, мышечной системы. Сюда же конвергируют сигналы от интерорецепторов зон проекции блуждающего и чревного нервов, гипоталамуса.
Нейроны неспецифических ядер образуют свои связи по ретикулярному типу. Их аксоны поднимаются в кору большого мозга и контактируют со всеми ее слоями, образуя не локальные, а диффузные связи. К неспецифическим ядрам поступают связи из ретикулярной формации ствола мозга, гипоталамуса, лимбической системы, базальных ганглиев, специфических ядер таламуса. Возбуждение неспецифических ядер вызывает генерацию в коре специфической веретенообразной электрической активности, свидетельствующей о развитии сонного состояния. Нарушение функции неспецифических ядер затрудняет появление веретенообразной активности, т. е. развитие сонного состояния. Сложное строение таламуса, наличие в нем взаимосвязанных специфических, неспецифических и ассоциативных ядер позволяет ему организовывать такие двигательные реакции, как сосание, жевание, глотание, смех. Двигательные реакции интегрируются в таламусе с вегетативными процессами, обеспечивающими эти движения.
Зрительный бугор является центром организации и реализации инстинктов, влечений, эмоций. Возможность получать информацию о состоянии множества систем организма позволяет таламусу участвовать в регуляции и определении функционального состояния организма в целом. Конвергенция сенсорных стимулов в таламус обусловливает возникновение так называемых таламических неукротимых болей, которые возникают при патологических процессах в самом таламусе.
В клинике симптомами поражения зрительных бугров являются сильная головная боль, расстройства сна, нарушения чувствительности, как в сторону повышения, так и понижения, нарушения движений, их точности, соразмерности, возникновение насильственных непроизвольных движений.
Гипоталамус (hypothalamus, подбугорье) – структура промежуточного мозга, входящая в лимбическую систему, организующая эмоциональные, поведенческие, гомеостатические реакции организма (рис. 25).
Гипоталамус имеет большое число нервных связей с корой большого мозга, подкорковыми узлами, зрительным бугром, средним мозгом, мостом, продолговатым и спинным мозгом.
 В состав гипоталамуса входят серый бугор, воронка с нейрогипофизом и сосцевидные тела. Морфологически в нейронных структурах гипоталамуса можно выделить около 50 пар ядер, имеющих свою специфическую функцию. Топографически эти ядра можно объединить в 5 групп: 1) передняя группа, в состав которой входят супраоптическое, паравентрикулярные ядра; 2) задняя группа сформирована из медиальных и латеральных ядер сосцевидных тел и заднего гипоталамического ядра; 3) средняя группа состоит из нижнемедиального и верхнемедиального ядер; 4) преоптическая группа имеет выраженные связи с конечным мозгом и делится на медиальное и латеральное предоптические ядра; 5) наружная группа включает в себя латеральное гипоталамическое поле и серобугорные ядра.
В состав гипоталамуса входят серый бугор, воронка с нейрогипофизом и сосцевидные тела. Морфологически в нейронных структурах гипоталамуса можно выделить около 50 пар ядер, имеющих свою специфическую функцию. Топографически эти ядра можно объединить в 5 групп: 1) передняя группа, в состав которой входят супраоптическое, паравентрикулярные ядра; 2) задняя группа сформирована из медиальных и латеральных ядер сосцевидных тел и заднего гипоталамического ядра; 3) средняя группа состоит из нижнемедиального и верхнемедиального ядер; 4) преоптическая группа имеет выраженные связи с конечным мозгом и делится на медиальное и латеральное предоптические ядра; 5) наружная группа включает в себя латеральное гипоталамическое поле и серобугорные ядра.
Ядра гипоталамуса имеют мощное кровоснабжение, подтверждением чему служит тот факт, что ряд ядер гипоталамуса обладает изолированным дублирующим кровоснабжением из сосудов артериального круга большого мозга (виллизиев круг). На 1 мм2 площади гипоталамуса приходится до 2600 капилляров, в то время как на той же площади V слоя предцентральной извилины (моторной коры) их 440, в гиппокампе – 350, в бледном шаре – 550, в затылочной доле коры большого мозга (зрительной коре) – 900. Капилляры гипоталамуса высокопроницаемы для крупномолекулярных белковых соединений, к которым относятся нуклеопротеиды, что объясняет высокую чувствительность гипоталамуса к нейровирусным инфекциям, интоксикациям, гуморальным сдвигам.
У человека гипоталамус окончательно созревает к 13 – 14 годам, когда заканчивается формирование гипоталамо-гипофизарных нейросекреторных связей. За счет мощных афферентных связей с обонятельным мозгом, базальными ганглиями, таламусом, гиппокампом, корой большого мозга гипоталамус получает информацию о состоянии практически всех структур мозга. В то же время гипоталамус посылает информацию к таламусу, ретикулярной формации, вегетативным центрам ствола мозга и спинного мозга.
Нейроны гипоталамуса имеют особенности, которые и определяют специфику функций самого гипоталамуса. К этим особенностям относятся чувствительность нейронов к составу омывающей их крови, отсутствие гематоэнцефалического барьера между нейронами и кровью, способность нейронов к нейросекреции пептидов, нейромедиаторов и др.
Влияние на симпатическую и парасимпатическую регуляцию позволяет гипоталамусу воздействовать на вегетативные функции организма гуморальным и нервным путями.
Раздражение ядер передней группы сопровождается парасимпатическими эффектами. Раздражение ядер задней группы вызывает симпатические эффекты в работе органов. Стимуляция ядер средней группы приводит к снижению влияний симпатического отдела вегетативной нервной системы. Указанное распределение функций гипоталамуса не абсолютно. Все структуры гипоталамуса способны в разной степени вызывать симпатические и парасимпатические эффекты. Следовательно, между структурами гипоталамуса существуют функциональные взаимодополняющие, взаимокомпенсирующие отношения.
В целом за счет большого количества связей, полифункциональности структур гипоталамус выполняет интегрирующую функцию вегетативной, соматической и эндокринной регуляции, что проявляется и в организации его ядрами ряда конкретных функций. Так, в гипоталамусе располагаются центры гомеостаза, теплорегуляции, голода и насыщения, жажды и ее удовлетворения, полового поведения, страха, ярости. Все эти центры реализуют свои функции путем активации или торможения автономного (вегетативного) отдела нервной системы, эндокринной системы, структур ствола и переднего мозга.
Особое место в функциях гипоталамуса занимает регуляция деятельности гипофиза. В гипоталамусе и гипофизе образуются также нейрорегуляторные пептиды – энкефалины, эндорфины, обладающие морфиноподобным действием и способствующие снижению стресса и т. д.
Нейроны ядер передней группы гипоталамуса продуцируют вазопрессин, или антидиуретический гормон (АДГ), окситоцин и другие пептиды, которые по аксонам попадают в заднюю долю гипофиза – нейрогипофиз.
Нейроны ядер срединной группы гипоталамуса продуцируют так называемые рилизинг-факторы, стимулирующие (либерины) и ингибирующие (статины) активность передней доли гипофиза – аденогипофиз. В нем образуются такие вещества, как соматотропный, тиреотропный и другие гормоны. Наличие такого набора пептидов в структурах гипоталамуса свидетельствует о присущей им нейросекреторной функции.
Нейроны гипоталамуса также обладают детектирующей функцией: реагируют на изменения температуры крови, электролитного состава и осмотического давления плазмы, количества и состав гормонов крови.
Исследования Дельгадо (Delgado) во время хирургических операций показали, что у человека раздражение определенных участков гипоталамуса вызывало эйфорию, эротические переживания. В клинике показано также, что патологические процессы в гипоталамусе могут сопровождаться ускорением полового созревания, нарушением менструального цикла, половой функции.
Раздражение передних отделов гипоталамуса может вызывать у животных пассивно-оборонительную реакцию, ярость страх, а раздражение заднего гипоталамуса вызывает активную агрессию. Кроме того, раздражение заднего гипоталамуса приводит к экзофтальму, расширению зрачков, повышению кровяного давления, сужению просвета артериальных сосудов, сокращениям желчного, мочевого пузырей. Могут возникать взрывы ярости с описанными симпатическими проявлениями. Уколы в области гипоталамуса вызывают глюкозурию, полиурию. В ряде случаев раздражение вызывало нарушение теплорегуляции: животные становились пойкилотермными, у них не возникало лихорадочное состояние.
Гипоталамус является также центром регуляции цикла бодрствование – сон. При этом задний гипоталамус активизирует бодрствование, стимуляция переднего вызывает сон. Повреждение заднего гипоталамуса может вызвать так называемый летаргический сон.
2. 3. 6 Конечный мозг (telencephalon).Конечный мозг является наиболее молодым в филогенетическом отношении. Он состоит из двух полушарий, каждое из которых представлено плащом, обонятельным мозгом и базальными или подкорковыми ганглиями (ядрами). Длина полушарий в среднем – 17 см, высота – 12 см. Полостью конечного мозга являются боковые желудочки, находящиеся в каждом из полушарий. Полушария большого мозга отделены друг от друга продольной щелью большого мозга и соединяются при помощи мозолистого тела, передней и задней спаек и спайки свода. Мозолистое тело состоит из поперечных волокон которые в латеральном направлении продолжаются в полушария, образуя лучистость мозолистого тела.
Обонятельный мозг представлен обонятельными луковицами, обонятельным бугорком, прозрачной перегородкой и прилежащими областями коры (препериформной, периамигдалярной и диагональной). Это самая меньшая часть конечного мозга, он обеспечивает функцию первого органа чувств, появившегося у живых существ, – функцию обоняния, и, кроме того, входит в состав лимбической системы. Повреждение структуры лимбической системы вызывает глубокое нарушение эмоций и памяти.
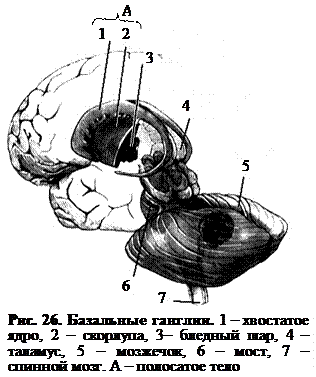 Базальные ганглии (ядра серого вещества) расположены в глубине больших полушарий. Они составляют примерно 3% от их объема. Базальные ганглии образуют многочисленные связи как между структурами, входящими в их состав, так и другими отделами мозга (корой больших полушарий, таламусом, черной субстанцией, красным ядром, мозжечком, мотонейронами спинного мозга). К базальным ганглиям относятся сильно вытянутое в длину и изогнутое хвостатое ядро (рис. 26.–1) и заложенное в толще белого вещества чечевицеобразное ядро. Двумя белыми пластинками оно подразделяется на наиболее крупную, лежащую латерально скорлупу и бледный шар (рис. 26.–2, 3). Хвостатое ядро и скорлупа объединяются под названием полосатого тела, они связаны анатомически и характеризуются чередованием белого и серого вещества.
Базальные ганглии (ядра серого вещества) расположены в глубине больших полушарий. Они составляют примерно 3% от их объема. Базальные ганглии образуют многочисленные связи как между структурами, входящими в их состав, так и другими отделами мозга (корой больших полушарий, таламусом, черной субстанцией, красным ядром, мозжечком, мотонейронами спинного мозга). К базальным ганглиям относятся сильно вытянутое в длину и изогнутое хвостатое ядро (рис. 26.–1) и заложенное в толще белого вещества чечевицеобразное ядро. Двумя белыми пластинками оно подразделяется на наиболее крупную, лежащую латерально скорлупу и бледный шар (рис. 26.–2, 3). Хвостатое ядро и скорлупа объединяются под названием полосатого тела, они связаны анатомически и характеризуются чередованием белого и серого вещества.
Полосатое тело принимает участие в организации и регуляции движений и обеспечении перехода одного вида движения в другое. Стимуляция хвостатого ядра тормозит восприятие зрительной, слуховой и других видов сенсорной информации, угнетает активность коры, подкорки, безусловные рефлексы (пищевой, оборонительный и др.) и выработку условных рефлексов, приводит к наступлению сна. При поражении полосатого тела наблюдается ретроантероградная амнезия – выпадение памяти на события, предшествующие травме. Двустороннее повреждение полосатого тела побуждает к стремлению движения вперед, одностороннее – приводит к манежным движениям. С нарушением функций полосатого тела связывают заболевание нервной системы – хорею (усиливаются вспомогательные и мимические движения). Скорлупа обеспечивает организацию пищевого поведения. При ее поражении наблюдаются трофические нарушения кожи, а ее раздражение вызывает слюноотделение и изменение дыхания.
Функции бледного шара заключаются в провоцировании ориентировочной реакции, движения конечностей, пищевого поведения (жевание, глотание).
После разрушения бледного шара возникают скованность движений, обеднение мимики (маскообразное лицо), гиподинамия, эмоциональная тупость, тремор головы, конечностей при движении, монотонная речь. При повреждениях бледного шара могут появиться подергивания отдельных мышц лица и туловища, нарушается синергизм движения конечностей при ходьбе. У человека с дисфункцией бледного шара затруднено начало движений, исчезают вспомогательные и реактивные движения при вставании, нарушаются содружественные движения рук при ходьбе, появляется симптом пропульсии: длительная подготовка к движению, затем быстрое движение и остановка. Такие циклы у больных повторяются многократно.
Плащ у человека представлен корой, т. е. пластинкой серого вещества, отделенной от полости желудочков белым веществом, которое содержит огромное количество нервных волокон, подразделяемых на три группы.
1. Ассоциативные волокна, или пути, соединяют различные отделы коры большого мозга внутри одного полушария. Выделяют короткие или дугообразные ассоциативные волокна, связывающие две лежащие рядом извилины, и длинные – протягивающиеся из одной доли в другую, оставаясь в пределах одного полушария.
2. Комиссуральные или спаечные волокна связывают кору обоих полушарий. Самой большой комиссурой головного мозга является мозолистое тело.
3. Проекционные пути связывают кору большого мозга с периферией. Существуют центробежные (эфферентные, двигательные) волокна, несущие нервные импульсы из коры на периферию, и центростремительные (афферентные, чувствительные) волокна, несущие импульсы с периферии в кору больших полушарий.
 Высшим отделом ЦНС является кора большого мозга (кора больших полушарий). Она обеспечивает совершенную организацию поведения животных на основе врожденных и приобретенных в онтогенезе функций.
Высшим отделом ЦНС является кора большого мозга (кора больших полушарий). Она обеспечивает совершенную организацию поведения животных на основе врожденных и приобретенных в онтогенезе функций.
Кора большого мозга делится на древнюю (archicortex), старую (paleocortex) и новую (neocortex). Древняя кора наряду с другими функциями имеет отношение к обонянию и обеспечению взаимодействия систем мозга. Старая кора включает поясную извилину, гиппокамп. У новой коры наибольшее развитие величины, дифференциации функций отмечается у человека. Толщина новой коры колеблется от 1,5 до 4,5 мм и максимальна в передней центральной извилине.
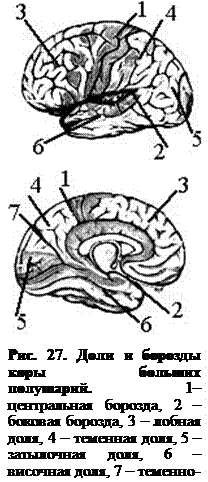
Кора полушарий покрыта бороздами и извилинами (рис. 27). Благодаря этому поверхность коры значительно увеличивается. Различают самые глубокие первичные борозды, которые делят полушария на доли. Боковая борозда (Сильвиева) отделяет лобную долю от височной, центральная борозда (Роландова) – лобную от теменной. Теменно-затылочная борозда располагается на медиальной поверхности полушария и разделяет теменную и затылочную доли, на верхнелатеральной поверхности явная граница между этими долями отсутствует. На медиальной поверхности располагается поясная борозда, переходящая в гиппокампову борозду, которые ограничивают обонятельный мозг от остальных долей.
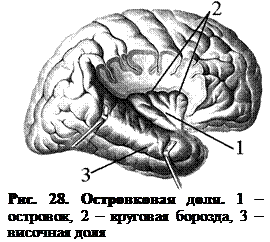
В глубине боковой борозды (рис. 28) располагается островковая доля. Она окружена с трех сторон круговой бороздой, ее поверхность изрезана бороздами и извилинами. Функционально островок связан с обонятельным мозгом.
 Вторичные борозды менее глубокие, они делят доли на извилины и располагаются снаружи от одноименных извилин. Третичные (безимянные) борозды придают извилинам индивидуальную форму, увеличивают площадь их коры.
Вторичные борозды менее глубокие, они делят доли на извилины и располагаются снаружи от одноименных извилин. Третичные (безимянные) борозды придают извилинам индивидуальную форму, увеличивают площадь их коры.
Функции отдельных зон новой коры определяются особенностями ее структурно-функциональной организации, связями с другими структурами мозга, участием в восприятии, хранении и воспроизведении информации при организации и реализации поведения, регуляции функций сенсорных систем, внутренних органов.
Особенности структурнофункцио- нальной организации коры большого мозга обусловлены тем, что в эволюции происходила кортикализация функций, т. е. передача коре большого мозга функций нижележащих структур мозга. Однако эта передача не означает, что кора берет на себя выполнение функций других структур. Ее роль сводится к коррекции возможных нарушений функций взаимодействующих с ней систем, более совершенного, с учетом индивидуального опыта, анализа сигналов и организации оптимальной реакции на эти сигналы, формирование в своих и в других заинтересованных структурах мозга памятных следов о сигнале, его характеристиках, значении и характере реакции на него. В дальнейшем, по мере автоматизации реакция начинает выполняться подкорковыми структурами.
Цитоархитектоника коры головного мозга. Общая площадь коры большого мозга человека около 2200 см2, число нейронов коры превышает 10 млрд. В составе коры имеются пирамидные, звездчатые, веретенообразные нейроны.
Пирамидные нейроны имеют разную величину, их дендриты несут большое количество шипиков; аксон пирамидного нейрона, как правило, идет через белое вещество в другие зоны коры или в структуры ЦНС.
Звездчатые клетки имеют короткие хорошо ветвящиеся дендриты и короткий аскон, обеспечивающий связи нейронов в пределах самой коры большого мозга.
Веретенообразные нейроны обеспечивают вертикальные или горизонтальные взаимосвязи нейронов разных слоев коры.
Кора большого мозга имеет преимущественно шестислойное строение (рис. 29).
 Слой I – верхний молекулярный, представлен в основном ветвлениями восходящих дендритов пирамидных нейронов, среди которых расположены редкие горизонтальные клетки и клетки-зерна, сюда же приходят волокна неспецифических ядер таламуса, регулирующие через дендриты этого слоя уровень возбудимости коры большого мозга.
Слой I – верхний молекулярный, представлен в основном ветвлениями восходящих дендритов пирамидных нейронов, среди которых расположены редкие горизонтальные клетки и клетки-зерна, сюда же приходят волокна неспецифических ядер таламуса, регулирующие через дендриты этого слоя уровень возбудимости коры большого мозга.
Слой II – наружный зернистый, состоит из звездчатых клеток, определяющих длительность циркулирования возбуждения в коре большого мозга, т. е. имеющих отношение к памяти.
Слой III – наружный пирамидный, формируется из пирамидных клеток малой величины и вместе со II слоем обеспечивают корко-корковые связи различных извилин мозга.
Слой IV – внутренний зернистый, содержит преимущественно звездчатые клетки. Здесь заканчиваются специфические таламокортикальные пути, т. е. пути, начинающиеся от рецепторов анализаторов.
Слой V – внутренний пирамидный (ганглиозный), слой крупных пирамид, которые являются выходными нейронами, аксоны их идут в ствол мозга и спинной мозг. В двигательной зоне в этом слое находятся гигантские пирамидные клетки, открытые Бецом (клетки Беца).
Слой VI – слой полиморфных клеток, большинство нейронов этого слоя образуют кортико-таламические пути.
Клеточный состав коры по разнообразию морфологии, функции, формам связи не имеет себе равных в других отделах ЦНС. Нейронный состав, распределение нейронов по слоям в разных областях коры различны, что позволило выделить в мозге человека 53 цитоархитектонических поля (поля Бродмана). Разделение коры большого мозга на цитоархитектонические поля более четко формируется по мере совершенствования ее функции в филогенезе.
У человека и высших млекопитающих различают, наряду с первичными, вторичные и третичные корковые поля, обеспечивающие ассоциацию функций данного анализатора с функциями других анализаторов. Для всех анализаторов характерен соматотопический принцип организации
Особенностью корковых полей является экранный принцип их функционирования. Этот принцип заключается в том, что рецептор проецирует свой сигнал не на один нейрон коры, а на поле нейронов, которое образуется их коллатералями и связями. В результате сигнал фокусируется не точка в точку, а на множестве разнообразных нейронов, что обеспечивает его полный анализ и возможность передачи в другие заинтересованные структуры. Так, одно волокно, приходящее в зрительную область коры, может активировать зону размером 0,1 мм. Это значит, что один аксон распределяет свое действие на более чем 5 000 нейронов.
Входные (афферентные) импульсы поступают в кору снизу, поднимаются к звездчатым и пирамидным клеткам III – V слоев коры. От звездчатых клеток IV слоя сигнал идет к пирамидным нейронам III слоя, а отсюда по ассоциативным волокнам – к другим полям, областям коры большого мозга. Звездчатые клетки поля 3 переключают сигналы, идущие в кору, на пирамидные нейроны V слоя, отсюда обработанный сигнал уходит из коры к другим структурам мозга.
В коре входные и выходные элементы вместе со звездчатыми клетками образуют так называемые колонки – функциональные единицы коры, организованные в вертикальном направлении. Доказательством этого служит следующее: если микроэлектрод погружать перпендикулярно в кору, то на своем пути он встречает нейроны, реагирующие на один вид раздражения, если же микроэлектрод вводить горизонтально по коре, то он встречает нейроны, реагирующие на разные виды стимулов.
Диаметр колонки около 500 мкм и определяется она зоной распределения коллатералей восходящего афферентного таламокортикального волокна. Соседние колонки имеют взаимосвязи, организующие участки множества колонок в организации той или иной реакции. Возбуждение одной из колонок приводит к торможению соседних.
Как уже упоминалось, разные области коры большого мозга имеют разные поля, определяющиеся по характеру и количеству нейронов, толщине слоев и т. д. Наличие структурно различных полей предполагает и разное их функциональное предназначение (рис. 30). Действительно, в коре большого мозга выделяют сенсорные, моторные и ассоциативные области.