III. Гидролазы.
К этому классу относятся ферменты, катализирующие процессы расщепления сложных органических веществ с помощью воды (гидролиз).
а) Полиазы. К ним относятся а-амилаза слюны, сока поджелудочной железы, крови и тканей животных (содержится в печени, мозге, мышцах). Она катализирует гидролиз крахмала и гликогена с образованием большого количества декстринов и немного мальтозы, причем действует только на 1,4 гликозидную связь.
б)Эстеразы. К эстеразам относятся ферменты, катализирующие гидролиз
сложных эфиров - это липазы, фосфатазы.
Липазы катализируют гидролиз жиров с образованием глицерина и жирных кислот, т.е. действие липаз направлено на сложноэфирную связь между Зх-атомным спиртом глицерином и жирной кислотой. Липазы содержатся в соке pancreas, печени, в сыворотке крови.
Фосфатазы катализируют гидролиз сложных эфиров, состоящих из спиртов и фосфорной кислоты. Среди фосфатаз различают специфически и неспецифически действующие. Специфически действующие гидролизуют определенные фосфорные эфиры, например, рибонуклеаза или дезоксирибонуклеаза гидролизуют полинуклеотиды с разрывом эфирной связи, идущей от остатков НзРО4 к спиртовой группе рибозы или дезоксирибозы соответственно. Неспецифически действующие фосфатазы катализируют отщепление НзРО4 от различных фосфорных эфиров независимо от входящих в их состав химической природы спиртов. Неспецифические фосфатазы разделяются на так называемые «кислые» и «щелочные». Оптимум действия первых при рН 4,5-50, вторых –при 8,5-9,0.
в) Протеазы (протеолитические ферменты). Они расщепляют белки и
полипептиды, гидролизуя пептидные связи.
Пепсин в виде профермента (пепсиногена) образуется в главных клетках слизистой оболочки желудка. Под действием НСI, образующейся в обкладочных клетках слизистой, пепсиноген превращается в пепсин. Пепсин катализирует гидролиз большинства белков, кроме протаминов, кератина волос и шерсти и др. Пепсин более интенсивно расщепляет денатурированные белки; кислая среда желудочного сока способствует денатурации белков, то есть действию пепсина.
Под влиянием пепсина начинается переваривание белков в желудке (до 30% их) с образованием в основном полипептидов.
Трипсин образуется в поджелудочной железе в виде профермента трипсиногена и в таком виде выделяется с соком этой железы. Активация трипсиногена (переход в трипсин) происходит под влиянием энтерокиназы - фермента, вырабатываемого в слизистой тонких кишок. Под влиянием трипсина белки расщепляются на полипептиды и аминокислоты.
Химотрипсин также образуется в pancreas в виде профермента химотрипсиногена. Активация последнего, то есть переход в химотрипсин происходит под влиянием ничтожно малых количеств трипсина. Химотрипсин тоже катализирует гидролиз белков и полипептидов.
Пептидазы гидролизуют полипептиды до аминокислот. Различают:
а) карбоксипептидаза, содержится в соке pancreas, отщепляет от пептида концевую аминокислоту, у которой имеется свободная карбоксильная группа.
б) аминопептидаза, образуется в слизистой кишечника и отщепляет от пептида концевую аминокислоту, у которой имеется свободная аминогруппа.
в) дипептидазы. Содержатся в слизистой кишечника, почках, лейкоцитах. Гидролизуют дипептид с образованием двух аминокислот.
IV. Лиазы. Это ферменты, катализирующие реакции негидролитического
расщепления молекул. При этом имеет место разрыв связи между атомами:
С-С, C-S, а также отщепление С02 от молекул органических веществ.
а) Альдолазы - это С-С-лиазы.

б) Декарбоксилазы- катализируют реакции отщепления С02 от
различных аминокислот с образованием аминов:

V.Изомеразы (мутазы). Это ферменты, катализирующие взаимопревращение субстратов. В качестве примера можно привести глюкозо-6-фосфат-изомеразу, катализирующую превращение глюкозо-6-фосфата во фруктозо-6-фосфат.
VI.Лигазы (синтетазы). К ним относятся ферменты, катализирующие реакции синтеза органических веществ с образованием химических связей С-С, C-S. Лигазы катализируют также присоединение отдельных аминокислот к соответствующим РНК (транспортным).
2 . 4 . ИЗОФЕРМЕНТЫ.
Это ферменты, катализирующие одну и ту же химическую реакцию, но отличающиеся между собой оптимумом рН, температуры, электролитической подвижностью, иммунохимическими свойствами, а также локализацией в тканях и набором субъединиц.
Например, лактатдегидрогеназа (ЛДГ) имеет 5 изоферментов, состоящих из «Н» (heart- сердце) и «М» (muscle- мышца) субъединиц:

ЛДГ1 локализуется преимущественно в мышце сердца, и повышение ее активности отмечается при патологии сердца (инфаркт миокарда). ЛДГ 2 и ЛДГ 3 повышают активность при заболеваниях крови. ЛДГ 4 и ЛДГ5 повышают активность при заболеваниях мышц;
ЛДГ 5 локализуется в ткани печени, и её активность повышается при заболеваниях печени, например, болезни Боткина.
Биологический смысл существования ЛДГ в виде изоферментов связан с тем, что ЛДГ1 но своим кинетическим параметрам более приспособлена для реакции лактат—►пируват, а ЛДГ5 - для пируват—►лактат. В разных метаболических ситуациях, возникающих в организме при различных нагрузках или заболеваниях, это позволяет органам и тканям легче приспосабливаться к новым условиям. Соответствующее отражение это находит и в сыворотке крови, где имеется соотношение (спектр) изоферментов ЛДГ.
Креатинкиназа (КФК) имеет 3 изофермента:
КФКММ – содержится в мышцах и повышается при их патологии;
КФКМБ –содержится в миокарде и повышается при заболеваниях сердца (инфаркт)
КФКББ - содержится в головном мозге
Биологическая роль изоферментов заключается также в обеспечении транспортных процессов между цитозолем и митохондриями. Для этого необходимы два изофермента того или иного фермента: цитозольный и митохондриальный. Такие процессы транспорта получили название челночных механизмов. Наибольшее значение имеют следующие из них:
1. Креатинфосфатный челночный механизм.

С помощью этого механизма осуществляется транспорт энергии АТФ из митихондрий в цитозоль.
2. Малатный челночный механизм:
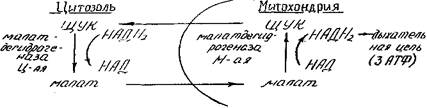
С помощью малатного механизма нейтрализуется избыток HAДH2 в цитозоле и осуществляется перенос восстанавливающих эквивалентов (протонов) из цитозоля в митохондрии, где их энергия трансформируется в АТФ.
Подобным же образом функционирует a - глицерофосфатный челночный механизм, где субстратами являются фосфодиоксиацетон и a - глицерофосфат, а изоферментами - цитозольная и митохондриальная формы глицерофосфат – дегидрогеназы.