Уровни регуляции активности ферментов.
Организм очень сложная система, и все процессы в нём в норме взаимосвязаны, без лишних реакций и напрасных затрат. Но т.к. организм не закрытая система, и постоянно испытывает воздействия извне, нужны механизмы регуляции, которые бы приспосабливали его к этим изменениям.
Поскольку всеми процессами в нашем организме управляют ферменты (гормоны действуют через фермент), то при изменении условий для соответствия процессов этим условиям будет меняться активность и количество ферментов.
1уровень. Изменения активности при изменении температуры, кол-ва субстрата, рН среды, т.к. в этих условиях меняется подвижность молекулы, ионизация функциональных групп, а следовательно, и активность фермента.
2уровень. Влияния активаторов и ингибиторов на работу фермента (его количество не меняется, меняется конформация) через аллостерич. и иногда активный центр.
3уровень. Индукция и репрессия синтеза Е, т.е. меняется его количество.
4уровень – организменный (нейрорегуляция). Происходит регуляция синтеза ферментов, участвующих в процессах нормализации гомеостаза. 4.1. гормональная – одни гормоны влияют на выделение других (релизинг-факторы: статины, либерины, а затем - тропные гормоны). 4.2. регуляция продукции гормона по типу обратной связи (почти всегда по отрицательной). 4.3. регуляция с участием структур ЦНС. 4.4. саморегуляция, зависит от параметров гомеостаза.(околощитовидная железа при
2+
снижении Са в крови увеличивает продукцию паратгормона).
Регуляции активности ферментов.
 1. Частичный протеолиз - активатор
1. Частичный протеолиз - активатор

 из неактивного фермента
из неактивного фермента
образуется активный. пептид
Это обеспечивает появление
активного фермента в нужный момент
и в нужном месте (пищеварительные ферменты; ферменты, участвующие в свёртывании крови).

 2. Белок – белковые C R
2. Белок – белковые C R

 взаимодействия в виде C R +4сАМР 2 R 4сАМР + 2 С
взаимодействия в виде C R +4сАМР 2 R 4сАМР + 2 С
присоединения или
отщепления регуляторных неактивн. ПК активная
субъединиц или регуляторов. Происходит связывание с АМР с регуляторн.
субъединицей (R) и тем самым освобождение
каталитической субъединицы, осуществляющей
фосфорилирование белков.
3. Фосфорилирование и
 дефосфорилирование – АТР АДР
дефосфорилирование – АТР АДР
основной механизм протеинкиназа

 контроля скорости белок ФП
контроля скорости белок ФП
р-ий. Н2О
 протеинфосфатаза
протеинфосфатаза
Р
Введение «-» заряженной фосфорной группы приводит к обратимым изменениям конформации, и к изменению активности фермента (гликогенсинтаза, тканевая липаза).
4. Аллостерическая:
*активатор взаимодействует
с аллостерич. центром à
изменяется конформ.à
 улучшение связывания S с Е
улучшение связывания S с Е
и скоростьреакции. фосфофруктокиназа инибируется АТФ
 * Ингитор взаимодействует изоцитратДГ игибируется АТФ,
* Ингитор взаимодействует изоцитратДГ игибируется АТФ,
с Е происходит ингибирование +
реакциив результате НАДН Н
невозможности
связывания S с Е.
Ингибирование ферментов бывает обратимое и необратимое (в таком случае образуется прочная ковалентная связь между S и Е)
ретроингибирование – ингибирование конечным продуктом при его избытке (метаболитами)
аллостерич.
 центр
центр



 А А
А А
S
 католич
католич
центр
 |  | ||

 У У
У У


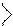 У
У
S
РИСУНОК на лист
Пояснение к рисунку.Клетка
* Белки в клетки проникают путём фаго- и пиноцитоза
 * АК (как в клетку, так и через систему внутриклеточных мембран) – путём активного транспорта (с затратой Е.) при участии - глутамил ТФ. и глутатиона.
* АК (как в клетку, так и через систему внутриклеточных мембран) – путём активного транспорта (с затратой Е.) при участии - глутамил ТФ. и глутатиона.
* Глю., вместе с Na+ – благодаря специфич. белку переносчику (вторичноактивный транспорт), который активируется с помощью инсулина (в инсулинзависимых клетках).
* Глицерин – свободно по градиенту концентраций.
* Ж.к. – свободно через плазматич. мембр., но их перемещение в матрикс МХ осуществляется с помощью карнитина.
* НАДН Н+, образующийся в результате гликолиза, не может проникать в МХ, челночные механизмы обеспечивают переброску 2Н с цитоплазматич. фракции НАДН Н+ на митохондриальную.
* образовавшийся в МХ ац.КоА не может пройти через МХ мембрану(в цитоплазме он необходим для синтеза ВЖК) , поэтому из него образуется цитрат, свободно через неё проникающий.
_
+СОО
 ПВ карбоксилаза о-ац
ПВ карбоксилаза о-ац
 ПВК
ПВК
ПВК-ДГ-ый комплекс ац.КоА
_
- СОО
Основные мембраны клетки и их функции