Типы экологических пирамид
1. пирамиды чисел - на каждом уровне откладывается численность отдельных организмов
Пирамида чисел отображает отчетливую закономерность, обнаруженную Элтоном: количество особей, составляющих последовательный ряд звеньев от продуцентов к консументам, неуклонно уменьшается (рис.3).
Например, чтобы прокормить одного волка, необходимо по крайней мере несколько зайцев, на которых он мог бы охотиться; чтобы прокормить этих зайцев, нужно довольно большое количество разнообразных растений. В данном случае пирамида будет иметь вид треугольника с широким основанием суживающимся кверху.
Однако подобная форма пирамиды чисел характерна не для всех экосистем. Иногда они могут быть обращенными, или перевернутыми. Это касается пищевых цепей леса, когда продуцентами служат деревья, а первичными консументами - насекомые. В этом случае уровень первичных консументов численно богаче уровня продуцентов (на одном дереве кормится большое количество насекомых), поэтому пирамиды чисел наименее информативны и наименее показательны, т.е. численность организмов одного трофического уровня в значительной степени зависит от их размеров.
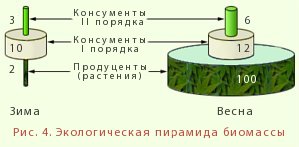
2. пирамиды биомасс - характеризует общую сухую или сырую массу организмов на данном трофическом уровне, например, в единицах массы на единицу площади - г/м2, кг/га, т/км2 или на объем - г/м3 (рис.4)
Обычно в наземных биоценозах общая масса продуцентов больше, чем каждого последующего звена. В свою очередь, общая масса консументов первого порядка больше, нежели консументов второго порядка и т.д.
В данном случае (если организмы не слишком различаются по размерам) пирамида также будет иметь вид треугольника с широким основанием суживающимся кверху. Однако и из этого правила имеются существенные исключения. Например, в морях биомасса растительноядного зоопланктона существенно (иногда в 2-3 раза) больше биомассы фитопланктона, представленного преимущественно одноклеточными водорослями. Это объясняется тем, что водоросли очень быстро выедаются зоопланктоном, но от полного выедания их предохраняет очень высокая скорость деления их клеток. В целом для наземных биогеоценозов, где продуценты крупные и живут сравнительно долго, характерны относительно устойчивые пирамиды с широким основанием. В водных же экосистемах, где продуценты невелики по размеру и имеют короткие жизненные циклы, пирамида биомасс может быть обращенной, или перевернутой (острием направлена вниз). Так, в озерах и морях масса растений превышает массу потребителей только в период цветения (весной), а в остальное время года может создаться обратное положение.
Пирамиды чисел и биомасс отражают статику системы, т. е. характеризуют количество или биомассу организмов в определенный промежуток времени. Они не дают полной информации о трофической структуре экосистемы, хотя позволяют решать ряд практических задач, особенно связанных с сохранением устойчивости экосистем.
Пирамида чисел позволяет, например, рассчитывать допустимую величину улова рыбы или отстрела животных в охотничий период без последствий для нормального их воспроизведения.

3. пирамиды энергии - показывает величину потока энергии или продуктивности на последовательных уровнях (рис.5).
В противоположность пирамидам чисел и биомассы, отражающим статику системы (количество организмов в данный момент), пирамида энергии отражая картину скоростей прохождения массы пищи (количества энергии) через каждый трофический уровень пищевой цепи, дает наиболее полное представление о функциональной организации сообществ.
На форму этой пирамиды не влияют изменения размеров и интенсивности метаболизма особей, и если учтены все источники энергии, то пирамида всегда будет иметь типичный вид с широким основанием и суживающейся верхушкой. При построении пирамиды энергии в ее основание часто добавляют прямоугольник, показывающий приток солнечной энергии.
Пирамиды энергии позволяют сравнивать энергетическую значимость популяций внутри экосистемы и иллюстрировать количественные отношения в отдельных, представляющих особый интерес частях экосистем, например, в звеньях жертва-хищник или хозяин-паразит.
В 1942 г. американский эколог Р. Линдеман сформулировал закон пирамиды энергий (закон 10 процентов), согласно которому с одного трофического уровня через пищевые цепи на другой трофический уровень переходит в среднем около 10% поступившей на предыдущий уровень экологической пирамиды энергии. Остальная часть энергии теряется в виде теплового излучения, на движение и т.д. Организмы в результате процессов обмена теряют в каждом звене пищевой цепи около 90% всей энергии, которая расходуется на поддержание их жизнедеятельности.
Если заяц съел 10 кг растительной массы, то его собственная масса может увеличиться на 1 кг. Лисица или волк, поедая 1 кг зайчатины, увеличивают свою массу уже только на 100 г. У древесных растений эта доля много ниже из-за того, что древесина плохо усваивается организмами. Для трав и морских водорослей эта величина значительно больше, поскольку у них отсутствуют трудноусвояемые ткани. Однако общая закономерность процесса передачи энергии остается: через верхние трофические уровни ее проходит значительно меньше, чем через нижние.
Рассмотрим превращение энергии в экосистеме на примере простой пастбищной трофической цепи, в которой имеется всего три трофических уровня.
1. уровень - травянистые растения,
2. уровень - травоядные млекопитающие, например, зайцы
3. уровень - хищные млекопитающие, например, лисы
Питательные вещества создаются в процессе фотосинтеза растениями, которые из неорганических веществ (вода, углекислый газ, минеральные соли и т.д.) с использованием энергии солнечного света образуют органические вещества и кислород, а также АТФ. Часть электромагнитной энергии солнечного излучения при этом переходит в энергию химических связей синтезируемых органических веществ.
Все органическое вещество, создаваемое в процессе фотосинтеза называется валовой первичной продукцией (ВПП). Часть энергии валовой первичной продукции расходуется на дыхание, в результате чего образуется чистая первичная продукция (ЧПП), которая и является тем самым веществом, которое поступает на второй трофический уровень и используется зайцами.
Пусть ВПП составляет 200 условных единиц энергии, а затраты растений на дыхание (R) - 50%, т.е. 100 условных единиц энергии. Тогда чистая первичная продукция будет равна: ЧПП = ВПП - R (100 = 200 - 100), т.е. на второй трофический уровень к зайцам поступит 100 условных единиц энергии.
Однако, в силу разных причин зайцы способны потребить лишь некоторую долю ЧПП (в противном случае исчезли бы ресурсы для развития живой материи), существенная же ее часть, в виде отмерших органических остатков (подземные части растений, твердая древесина стеблей, ветвей и т.д.) не способна поедаться зайцами. Она поступает в детритные пищевые цепи и (или) подвергается разложению редуцентами (F). Другая часть идет на построение новых клеток (численность популяции, прирост зайцев - Р) и обеспечение энергетического обмена или дыхания (R).
В этом случае, согласно балансовому подходу, балансовое равенство расхода энергии (С) будет выглядеть следующим образом: С = Р + R + F, т.е. поступившая на второй трофический уровень энергия будет израсходована, согласно закону Линдемана, на прирост популяции - Р - 10%, остальные 90% будут израсходованы на дыхание и удаление неусвоенной пищи.
Таким образом, в экосистемах с повышением трофического уровня происходит быстрое уменьшение энергии, накапливаемой в телах живых организмов. Отсюда ясно почему каждый последующий уровень всегда будет меньше предыдущего и почему цепи питания обычно не могут иметь более 3-5 (редко 6) звеньев, а экологические пирамиды не могут состоять из большого количества этажей: к конечному звену пищевой цепи так же, как и к верхнему этажу экологической пирамиды, будет поступать так мало энергии, что ее не хватит в случае увеличения числа организмов.
Такая последовательность и соподчиненность связанных в форме трофических уровней групп организмов представляет собой потоки вещества и энергии в биогеоценозе, основу его функциональной организации.
Важнейший вид взаимоотношений между организмами в биоценозе, фактически формирующими его структуру, – это пищевые связи хищника и жертвы: одни – поедающие, другие – поедаемые. При этом все организмы, живые и мертвые, являются пищей для других организмов: заяц ест траву, лиса и волк охотятся на зайцев, хищные птицы (ястребы, орлы и т.п.) способны утащить и съесть как лисенка, так и волчонка. Погибшие растения, зайцы, лисы, волки, птицы становятся пищей для детритофагов (редуцентов или иначе деструкторов).
Пищевая цепь – это последовательность организмов, в которой каждый из них съедает или разлагает другой. Она представляет собой путь движущегося через живые организмы однонаправленного потока поглощенной при фотосинтезе малой части высокоэффективной солнечной энергии, поступившей на Землю. В конечном итоге эта цепь возвращается в окружающую природную среду в виде низкоэффективной тепловой энергии. По ней также движутся питательные вещества от продуцентов к консументам и далее к редуцентам, а затем обратно к продуцентам.
Каждое звено пищевой цепи называют трофическим уровнем. Первый трофический уровень занимают автотрофы, иначе именуемые первичными продуцентами. Организмы второго трофического уровня называют первичными консументами, третьего – вторичными консументами и т.д. Обычно бывают четыре или пять трофических уровней и редко более шести (рис. 1)[1].
Существуют два главных типа пищевых цепей – пастбищные (или «выедания») и детритные (или «разложения»).
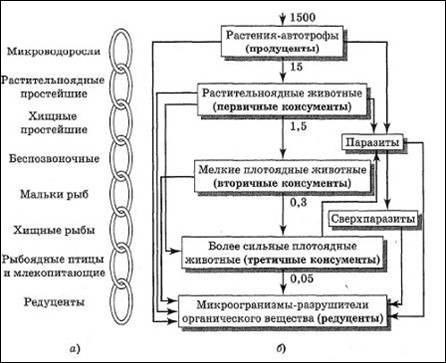
Рис. 1. Пищевые цепи биоценоза по Н.Ф. Реймерсу: обобщенная (а) и реальная (б)
Стрелками на рисунке 1 показано направление перемещения энергии, а цифрами – относительное количество энергии, приходящей на трофический уровень.
В пастбищных пищевых цепях первый трофический уровень занимают зеленые растения, второй – пастбищные животные (термин «пастбищные» охватывает все организмы, питающиеся растениями), а третий – хищники. Так, пастбищными пищевыми цепями являются:
РАСТИТЕЛЬНЫЙ МАТЕРИАЛ (например, нектар) => МУХА => ПАУК =>
=> ЗЕМЛЕРОЙКА => СОВА
СОК РОЗОВОГО КУСТА => ТЛЯ => БОЖЬЯ КОРОВКА => ПАУК =>
=> НАСЕКОМОЯДНАЯ ПТИЦА => ХИЩНАЯ ПТИЦА.
Детритная пищевая цепь начинается с детрита по схеме:
ДЕТРИТ-> ДЕТРИТОФАГ -> ХИЩНИК
Характерными детритными пищевыми цепями являются:
ЛИСТОВАЯ ПОДСТИЛКА ЛЕСА => ДОЖДЕВОЙ ЧЕРВЬ => ЧЕРНЫЙ ДРОЗД =>
=> ЯСТРЕБ-ПЕРЕПЕЛЯТНИК
МЕРТВОЕ ЖИВОТНОЕ => ЛИЧИНКИ ПАДАЛЬНОЙ МУХИ => ТРАВЯНАЯ ЛЯГУШКА => ОБЫКНОВЕННЫЙ УЖ.
Концепция пищевых цепей позволяет в дальнейшем проследить круговорот химических элементов в природе, хотя простые пищевые цепи, подобные изображенным ранее, где каждый организм представлен как питающийся организмами только какого-то одного типа, в природе встречаются редко.
Реальные пищевые связи намного сложнее, ибо животное может питаться организмами разных типов, входящих в одну и ту же пищевую цепь или в различные цепи, что особенно характерно для хищников (консументов) высших трофических уровней. Связь между пастбищной и детритной пищевыми цепями иллюстрирует предложенная Ю. Одумом модель потока энергии (рис. 2)[2].
Всеядные животные (в частности, человек) питаются и консументами, и продуцентами. Таким образом, в природе пищевые цепи переплетаются, образуют пищевые (трофические) сети.

Рис. 2. Схема пастбищной и детритнои пищевых цепей (по Ю. Одуму)
Правило Линдемана (10%)
Сквозной поток энергии, проходя через трофические уровни биоценоза, постепенно гасится. В 1942 г. Р. Линдеман сформулировал закон пирамиды энергий, или закон (правило) 10%, согласно которому с одного трофического уровня экологической пирамиды переходит на другой, более высокий ее уровень (по «лестнице»: продуцент – консумент – редуцент) в среднем около 10% поступившей на предыдущий уровень экологической пирамиды энергии. Обратный поток, связанный с потреблением веществ и продуцируемой верхним уровнем экологической пирамиды энергии более низкими ее уровнями, например, от животных к растениям, намного слабее – не более 0,5% (даже 0,25%) от общего ее потока, и потому говорить о круговороте энергии в биоценозе не приходится.
Если энергия при переходе на более высокий уровень экологической пирамиды десятикратно теряется, то накопление ряда веществ, в том числе токсичных и радиоактивных, в примерно такой же пропорции увеличивается. Этот факт фиксирован в правиле биологического усиления. Оно справедливо для всех ценозов. В водных биоценозах накопление многих токсичных веществ, в том числе хлорорганических пестицидов, коррелирует с массой жиров (липидов), т.е. явно имеет энергетическую подоснову.
45.
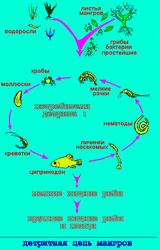
Пищевые цепи можно разделить на два типа. Пастбищная цепь начинается от зеленого растения и идет далее к пасущимся растительноядным животным и затем - к хищникам. Примеры пастбищных цепей приведены на иллюстрациях к параграфу 4.2. Детритная цепь идет от мертвого органического вещества (детрита) к микроорганизмам-редуцентам и животным, поедающим мертвые остатки (детритофагам), и затем - к хищникам, питающимся этими животными и микробами. На этом рисунке показан пример детритной пищевой цепи из тропиков; это цепь, начинающаяся от опадающих листьев мангров - деревьев и кустарников, растущих на периодически затопляемых приливами морских побережьях и в устьях рек. Их листья падают в солоноватые воды, заросшие мангровыми деревьями, и разносятся течением по обширной площади заливов. В воде на опавших листьях развиваются грибы, бактерии и простейшие, которых вместе с листьями поедают многочисленные организмы: рыбы, моллюски, крабы, рачки, личинки насекомых и круглые черви - нематоды. Этими животными питаются мелкие рыбы (например, гольяны), а их в свою очередь, поедает крупная рыба и хищные рыбоядные птицы. |
ПИЩЕВАЯ ЦЕПЬ (трофическая цепь, цепь питания), взаимосвязь организмов через отношения пища – потребитель (одни служат пищей для других). При этом происходит трансформация вещества и энергии от продуцентов(первичных производителей) через консументов (потребителей) к редуцентам (преобразователям мёртвой органики в неорганические вещества, усваиваемые продуцентами).
Различают 2 типа пищевых цепей – пастбищную и детритную. Пастбищная цепь начинается с зелёных растений, идёт к пасущимся растительноядным животным (консументы 1-го порядка) и затем к хищникам, добывающим этих животных (в зависимости от места в цепи – консументы 2-го и последующих порядков). Детритная цепь начинается с детрита (продукт распада органики), идёт к микроорганизмам, которые им питаются, а затем к детритофагам (животные и микроорганизмы, вовлечённые в процесс разложения отмирающей органики).
Примером пастбищной цепи может служить многоканальная её модель в африканской саванне. Первичными продуцентами являются травостой и деревья, консументами 1-го порядка – растительноядные насекомые и травоядные животные (копытные, слоны, носороги и др.), 2-го порядка – хищные насекомые, 3-го – плотоядные пресмыкающиеся (змеи и др.), 4-го – хищные млекопитающие и хищные птицы. В свою очередь детритофаги (жуки-скарабеи, гиены, шакалы, грифы и т. д.) на каждом из этапов пастбищной цепи разрушают туши погибших животных и остатки пищи хищников. Количество особей, включённых в пищевую цепь, в каждом её звене последовательно уменьшается (правило экологической пирамиды), т. е. число жертв всякий раз существенно превышает число их потребителей. Пищевые цепи не изолированы одна от другой, а переплетаются друг с другом, образуя пищевые сети.

44.
ддержание жизнедеятельности организмов и круговорот вещества в экосистемах, т. е. существование экосистем, зависит от постоянного притока энергии, необходимой всем организмам для их жизнедеятельности и самовоспроизведения (рис. 12.19).

Рис. 12.19. Поток энергии в экосистеме (по Ф. Рамаду, 1981)
В отличие от веществ, непрерывно циркулирующих по разным блокам экосистемы, которые всегда могут повторно использоваться, входить в круговорот, энергия может быть использована только раз, т. е. имеет место линейный поток энергии через экосистему.
Одностороний приток энергии как универсальное явление природы происходит в результате действия законов термодинамики. Первый закон гласит, что энергия может превращаться из одной формы (например, света) в другую (например, потенциальную энергию пищи), но не может быть создана или уничтожена. Второй закон утверждает, что не может быть ни одного процесса, связанного с превращением энергии, без потерь некоторой ее части. Определенное количество энергии в таких превращениях рассеивается в недоступную тепловую энергию, а следовательно, теряется. Отсюда не может быть превращений, к примеру, пищевых веществ в вещество, из которого состоит тело организма, идущих со 100-процентной эффективностью.
Таким образом, живые организмы являются преобразователями энергии. И каждый раз, когда происходит превращение энергии, часть ее теряется в виде тепла. В конечном итоге вся энергия, поступающая в биотический круговорот экосистемы, рассеивается в виде тепла. Живые организмы фактически не используют тепло как источник энергии для совершения работы — они используют свет и химическую энергию.
Пищевые цепи и сети, трофические уровни. Внутри экосистемы содержащие энергию вещества создаются автотрофными организмами и служат пищей для гетеротрофов. Пищевые связи — это механизмы передачи энергии от одного организма к другому.
Типичный пример: животное поедает растения. Это животное, в свою очередь, может быть съедено другим животным. Таким путем может происходить перенос энергии через ряд организмов — каждый последующий питается предыдущим, поставляющим ему сырье и энергию (рис. 12.20).
Рис. 12.20. Биотический круговорот веществ: пищевая цепь
(по А. Г. Банникову и др., 1985)
Такая последовательность переноса энергии называется пищевой (трофической) цепью,или цепью питания. Место каждого звена в цепи питания является трофическим уровнем.Первый трофический уровень, как уже было отмечено ранее, занимают автотрофы, или так называемые первичные проду-з центы. Организмы второгого трофического уровня называются первичными консументами, третьего — вторичными консументами и т. д.
Обычно различают три типа пищевых цепей. Пищевая цепь хищников начинается с растений и переходит от мелких организмов к организмам все более крупных размеров. На суше пищевые цепи состоят из трех-четырех звеньев.
Одна из простейших пищевых цепей имеет вид (см. рис. 12.5):
растение ® заяц ® волк
продуцент ® травоядное ® плотоядное
Широко распространены и такие пищевые цепи:
растительный материал (например, нектар) ® муха ® паук ®
землеройка ® сова.
сок розового куста ® тля ® божья (тлевая) коровка ®
® паук ® насекомоядная птица ® хищная птица.

43.
Различия между экосистемой и биогеоценозом можно свести к следующим положениям:
1) биогеоценоз - понятие территориальное, относится к конкретным участкам суши и имеет определенные границы, совпадающие с границами фитоценоза. Характерная особенность биогеоценоза, на которую указывают Н.В. Тимофеев-Ресовский, А.Н. Тюрюканов (1966) – через территорию биогеоценоза не проходит ни одна существенная биоценотическая, почвенно-геохимическая, геоморфологическая и микроклиматическая граница.
- понятие экосистемы шире, чем понятие биогеоценоза; оно применимо к биологическим системам разной сложности и размеров; экосистемы часто не имеют определенного объема и строгих границ;
2) в биогеоценозе органическое вещество всегда продуцируют растения, поэтому основной компонент биогеоценоза – фитоценоз;
- в экосистемах органическое вещество не всегда создается живыми организмами, нередко поступает извне.
(приносится течением – озеро, море; вносится человеком – сельскохозяйственные угодья, переносится ветром или осадками – растительные остатки на эродированных склонах гор).
3) биогеоценоз потенциально бессмертен;
- существование экосистемы может закончиться с прекращением прихода в нее вещества или энергии.
4) экосистема может быть и наземным и водным образованием;
- биогеоценоз всегда наземная или мелководная экосистема.
5) – в биогеоценозе всегда должен быть единый эдификатор (эдификаторная группировка или синузия), определяющий всю жизнь и строй системы.
- В экосистеме их может быть несколько.
На ранних стадиях развития экосистема склона – это будущий лесной ценоз. Она состоит из группировок организмов с разными эдификаторами и довольно неоднородными условиями среды. Лишь в будущем на одну и ту же группировку могут оказывать влияние не только её эдификатор, но и эдификатор ценоза. И второй будет основным.
Таким образом, не каждая экосистема является биогеоценозом, но каждый биогеоценоз – экосистема, полностью соответствующая определению Тенсли.
Различия между экосистемой и биогеоценозом можно свести к следующим положениям:
1) биогеоценоз - понятие территориальное, относится к конкретным участкам суши и имеет определенные границы, совпадающие с границами фитоценоза. Характерная особенность биогеоценоза, на которую указывают Н.В. Тимофеев-Ресовский, А.Н. Тюрюканов (1966) – через территорию биогеоценоза не проходит ни одна существенная биоценотическая, почвенно-геохимическая, геоморфологическая и микроклиматическая граница.
- понятие экосистемы шире, чем понятие биогеоценоза; оно применимо к биологическим системам разной сложности и размеров; экосистемы часто не имеют определенного объема и строгих границ;
2) в биогеоценозе органическое вещество всегда продуцируют растения, поэтому основной компонент биогеоценоза – фитоценоз;
- в экосистемах органическое вещество не всегда создается живыми организмами, нередко поступает извне.
(приносится течением – озеро, море; вносится человеком – сельскохозяйственные угодья, переносится ветром или осадками – растительные остатки на эродированных склонах гор).
3) биогеоценоз потенциально бессмертен;
- существование экосистемы может закончиться с прекращением прихода в нее вещества или энергии.
4) экосистема может быть и наземным и водным образованием;
- биогеоценоз всегда наземная или мелководная экосистема.
5) – в биогеоценозе всегда должен быть единый эдификатор (эдификаторная группировка или синузия), определяющий всю жизнь и строй системы.
- В экосистеме их может быть несколько.
На ранних стадиях развития экосистема склона – это будущий лесной ценоз. Она состоит из группировок организмов с разными эдификаторами и довольно неоднородными условиями среды. Лишь в будущем на одну и ту же группировку могут оказывать влияние не только её эдификатор, но и эдификатор ценоза. И второй будет основным.
Таким образом, не каждая экосистема является биогеоценозом, но каждый биогеоценоз – экосистема, полностью соответствующая определению Тенсли.
Экологическая структура биогеоценоза. Каждый биогеоценоз слагается из определенных экологических групп организмов, соотношение которых отражает экологическую структуру сообщества, складывающуюся в течение длительного времени в определенных климатических, почвенно-грунтовых и ландшафтных условиях строго закономерно. Например, в биогеоценозах разных природных зон закономерно изменяется соотношение фитофагов (животных, питающихся растениями) и сапрофагов. В степных, полупустынных и пустынных районах фитофаги преобладают над сапрофагами, а в лесных сообществах, наоборот, сильнее развита сапрофагия. В глубинах океана основным типом питания является хищничество, тогда как на освещенной поверхности водоема преобладают фильтраторы, потребляющие фитопланктон, либо виды со смешанным питанием.
42..