Отдел Риниеобразные (Риниофиты) – Rhyniophyta
К отделу принадлежат самые древние из известных высших растений (рис. 2). Названия рода Риния (Rhynia) происходит от деревни Райни (недалеко от г. Абердина) в Шотландии, близ которой были обнаружены остатки этих примитивных растений в кремнистых породах врачом У. Макки в 1912 г. Эти отложения представляют собой пропитанные растворами кремниевой кислоты окаменевшие девонские торфа. У. Макки в свободное от медицинской практики время увлекался геологией. Свои материалы по ископаемым таксонам он передал шотландскому палеоботанику Р. Кидстону, который совместно с профессором Манчестерского университета У. Лангом опубликовали в 1917–1921 гг. результаты изучения растений ископаемой флоры. Они заложили начало систематическому изучению группы древнейших и наиболее примитивных высших растений.

 ан
ан


 фт ст с арх
фт ст с арх

 м
м
3б
3а 4

1 5



р рз 8
 7б
7б



7а 9 10 11
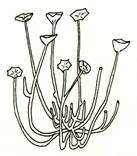
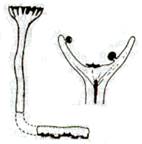
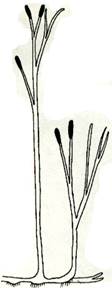
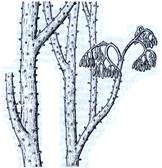
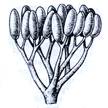
Рис. 2. Риниеобразные (Rhyniophyta): 1 – риния большая (Rhynia major), (м – мезомы, р – ризоиды, рз – ризомоид, ст – стерильные теломы, фт – фертильные теломы); 2 – хорнеофитон (Horneophyton), (с – спорангии); 3а,3б – лионофитон (Lyonophyton), (3а – реконструкция гаметофита; 3б – обоеполый гаметангиофор: ан – антеридий, арх – архегоний); 4 – сциадофитон (Sciadophyton) с однополыми гаметангиофорами; 5 – спорангии хедеи (Hedeia); – синангий ярравии (Jarravia); 7а,7б – пертика (Pertica), (7а – реконструкция спорофита, 7б – сильно разветвленные спороносные оси); 8 – фрагмент псилофита первичного (Psilophyton princeps); 9 – зостеролист (Zosterophyllum); 10 – госслингия (Gosslingia); 11 – савдония (Sawdonia).
Открытие У. Макки является вторым. Пальма первенства принадлежит канадскому геологу Джеймсу Досону, обнаружившему в 1859 г. в девонских отложениях на полуострове Гаспе (юго-восток Канады) остатки удивительно примитивного растения. Ему было дано название Psilophyton princeps («голорос первичный»). Отсюда и более раннее название этой группы растений – Псилофиты (Psilophyta). Растение представляло собой систему вильчато разветвленных осей, покрытых небольшими шипиками. На окончаниях дуговидно изогнутых поникающих веточек располагались спорангии. Псилофит не был похож ни на одно из растений, известных в то время науке и не укладывался в существовавшие тогда системы классификаций. Ценная находка Д. Досона долгое время оставалась почти незамеченной для науки загадкой природы. Положение изменилось только в начале ХХ века, когда они были открыты вновь У. Макки. Риния отличалась от голороса отсутствием выростов на разветвленных осях и вертикально расположенными конечными спорангиями. В 1937 г. У. Лангом в силурийских песчаниках Шотландии была обнаружена одна из наиболее древних палеоботанических находок высших растений – куксония (Cooksonia), обитавшая на Земле более 415 млн. лет назад. Она представляла собой похожий на водоросль «кустик» разветвленных уплощенных зеленых голых осей, несущих шаровидные спорангии. Прикреплялись растения к субстрату с помощью ризоидов. Это было третье крупное открытие наиболее примитивных высших растений. Полагают, что обнаруженные растения были представителями уже достаточно далеко разошедшихся в процессе быстрой эволюции таксономических групп. Это подтвердили более поздние находки новых ископаемых растений, их детальное изучение. Вероятно, первенцы наземной флоры имели еще более простое строение.
Древнейшие наземные растения раньше объединяли в один таксон под названием Псилофиты. Теперь разными исследователями предлагается 3 таксона на уровнях отделов. Многие объединяют их в отдел Риниеобразные – Rhyniophyta, насчитывающий около 20 родов и большое число видов. Единство их подтверждается рядом важных общих признаков, отражающих начальную ступень в эволюции высших растений. Единство их подкрепляемся также наличием родов, которые сочетают признаки трех отмеченных групп.
Отдел включает самые древние достоверные и по мнению большинства исследователей – самые примитивные травянистые высшие растения. Они появились приблизительно 430 млн. лет назад в силурийском периоде палеозойской эры. Геологическая история отдела заканчивается в девоне (около 370 млн. лет назад). Они дали начало другим группам высших растений.
Для них характерны черты крайне примитивного строения спорофита, диминировавшего в жизненном цикле. Они еще не имели типичных для высших растений олиственных побегов, корней и даже листоподобных придатков. Из-за отсутствия листьев эта группа растений была названа «голоростовые». Некоторые представители внешне были скорее похожи на водоросли, чем на высшие растения. Изучение их особенностей имело огромное значение для выяснения исходной модели строения высших растений и разработки направлений их морфологической эволюции. В конце Х1Х и начале ХХ веков было передоложено несколько гипотез, однако большинство современных ботаников придерживаются телломной теории, в разработке которой вклад внес известный немецкий ботаник В. Циммерман (30–40-е гг. ХХ века). Её основы были заложены ещё в трудах Г. Потонье, О. Линье, Ф. Боуэра. Большой вклад в развитие и становление теломной теории внес академик А.Л. Тахтаджян, ранее – Л.М. Кречетович, Б.М. Козо-Полянский и др.
Вегетативное тело спорофита риниофотов представлено безлистными цилиндрическими дихотомически ветвящимися, первично васкуляризованными осевыми структурами. Наземные оси выполняли в первую очередь функцию воздушного питания. Их конечные ответвления – теломы (от греч. telos – конец), а соединяющие их части, расположенные между точками ветвления – мезомы (от греч. mesos – средний). Одни теломы были стерильными, другие – фертильными и заканчивались спорангиями (рис 2). Стерильные теломы осуществляли только функцию фотосинтеза. Горизонтально расположенные на поверхности или в почве влажных местообитаний теломы получили название ризомоиды (т. е. корневищеподобные). От них отходили многочисленные простые (несептированные) одноклеточные ризоиды, осуществляя функцию минерального питания. Ризомоид – прототип корня, ризоиды – прототипы корневых волосков. Наличием проводящих пучков теломы принципиально отличались от структур водорослей.
Таким образом, теломы, мезомы и ризомоиды были первыми конструктивными элементами спорофита наземных растений. Возникновение этих элементарных органов было крупным ароморфозом в эволюции растительного мира. Это сильно повысило уровень организации растений, увеличило их пластичность и открыло более широкие возможности дальнейшего эволюционного развития.
Ветвление надземных вертикальных осей спорофита риниофитов характеризовалось равной и неравной дихотомией. Вследствие резко выраженной анизофилии, ветвление приобретало характер псевдомоноподиального. Поверхность надземных осей была голой или покрыта многочисленными выростами в виде энаций (в т.ч. шипиками, зубчиками).
Риниофиты – первые растения суши. Как наземные растения, в отличие от водорослей, они были покрыты типичной эпидермой, характеризовались наличием проводящей системы. Наружные стенки клеток эпидермы утолщены и покрыты слоем кутикулы. Имелись немногочисленные просто устроенные устьица (доказательство их наземного существования) – с двумя замыкающими клетками и устричной щелью. Они были лишены побочных клеток. Наличие устьиц в первую очередь отличает эпидерму высшего растения от наружного слоя клеток многоклеточных талломов (слоевищ) водорослей. Проводящая система слабо развита и построена по типу гаплостели, которая была началом развития всех остальных типов стели. Ксилема состояло из трахеид с кольчатыми, спиральными или реже лестничными утолщениями. Риниофты не обладали способностью к вторичному росту (камбий отсутствовал, как и перицикл). Меристема только верхушечная. В отличие от большинства сосудистых растений отсутствовали также и механические (опорные) ткани. Их функции выполняла толстая кора.
Спорангии всегда непосредственно прикреплялись к осям, или являлись их продолжением. У большинства более высоко организованных высших растений они прикрепляются к специализированным придаткам.
Спорангии риниофитов располагались верхушечно (терминально) на теломах или в боковом положении, одиночно (риния, куксония, хорнеофитон – Horneophyton, госслингия – Gosslingia) или группами (пертика – Pertica, тримерофит – Trimerophyton, зостеролист – Zosterophyllum). Они также отличались достаточно примитивным строением. Имели толстую многослойную стенку (спорангии эуспорангиатного типа), споры многочисленные, одинакового строения (растения морфологически равноспоровые). Спорангии различались по величине и форме. У наиболее примитивных видов они мало отличались от верхушек вегетативных осей. У куксонии они шаровидные, диаметром 1 мм. У ринии – продолговато-цилиндрические, длиной до 12 мм и толщиной до 4 мм. Спорангии не имели специальных приспособлений для вскрывания или они были крайне примитивно плана. У одних видов споры освобождались после разрушения стенки спорангия. У других видов спорангии вскрывались трещиной, располагавшейся продольно, поперечно (близ основания или верхушки спорангия), апикальной порой (у хорнеофитона). Нередко трещины проходили по определенным бороздам. В спорангий хорнеофитона глубоко вдавалась колонка, на которой располагался куполообразный спорангий. Интересно, но уже среди риниофитов наблюдалась тенденция к сближению спорангиев (хедея – Hedeia) и в дальнейшем к их срастанию в синангии (ярравия – Jarravia). Ярравия – нижнедевонское растение. В дальнейшем синангии будут неоднократно формироваться в различных линиях эволюции высших растений. Таким образом, спороносные структуры риниофитов, несмотря на примитивность их организации, характеризуются разнообразием морфологического строения. Эта особенность характерна для представителей большинства отделов высших растений и широко используется при классификации.
В спорангиях развивались многочисленные споры, одинакового морфологического строения (риниофиты были равноспоровыми растениями). Строение толстой оболочки спор – типичное для высших растений. Они были снабжены трехлучевым тетрадным рубцом.
Гаметофиты ринофитов существовали независимо от спорфитов и были свободно живущими растениями, однодомными (антеридии и архегонии развивались на одном растении). Под родовым названием лионофитон (Lyonophyton) описаны гаметофиты с обоеполыми гаметангиофорами. Предполагается, что они принадлежали хорнеофитону. На вершине некоторых неразветвленных осей имелось апикальное чашевидное расширение с лопастным краем. В центре чаши формировался небольшой выступ (архегониефор), к которому прикреплялись архегонии с сильно вытянутыми шейками. Булавовидные антеридии располагались по периферии чаши на внутренних стенках. Выявлены также гаметофиты с раздельнополыми гамитангиофорами (сциадофитон – Sciadophyton).
Произрастали риниофиты во влажных и болотистых местообитаниях вокруг морских и материковых водных бассейнов, а также в прибрежных мелководьях. Иногда они образовывали значительные заросли, являлись ландшафтными растениями. Возможно среди них были сухопутные растения. Некоторые были водными растениями с выступающими над водой спороносными частями.
Наиболее древней из достоверных находок высших растений является куксония. Более ранние риниофиты описаны только по спорам. Уже древнейшие силурийские риниофиты прошли длительный путь эволюционного развития. Это были довольно сложными, специализированными растениями разнообразного строения, приспособленными к жизни в наземных условиях.
Произошли они от зеленых водорослей. Наиболее древние формы еще сохраняли сходство с водорослеообразными предками. В частности, очень сходный характер их ветвления.
Многообразие признаков риниофитов явилось основой для дальнейших эволюционных направлений. Они дали начало многим независимым группам высших растений. В первую очередь плунообразным, хвощеобразным, псилотообразным и папоротникообразным, а также мохообразным (одна из гипотез). Изучение строения первенцев наземной суши имеет большое значение для эволюционной морфологии и филогении всего подцарства высших растений. Установлено, что первым органом спорофита высших растений был дихотомически разветвленный радиальный телом, строение которого, как отмечено, характерно для сухопутных растений (наличие проводящей системы, появление эпидермы, устьиц и т. д.). В ходе дальнейшего приспособления к наземной жизни от них произошли растения, обладавшие побегами и корнями.
Теломы риниофотов были цилиндрическими и имели косо-вертикальную ориентацию. К солнечным лучам была обращена только небольшая часть их поверхности. Увеличение размеров спорофита с целью улучшения рассеивания спор, требовало увеличения размеров поверхности воспринимающей свет. Это достигалось образованием уплощенных органов – листьев, ориентированных более горизонтально. Осевые органы, несущие листья, превращались в стебли. Постепенно возникли листостебельные растения. Листья возникали двумя способами (по пути «энационной микрофиллии» и «кладофильной макрофиллии»). У ископаемых плаунообразных многочисленные цельные мелкие листья (микрофиллы) возникают как боковые выросты (энации) на осевых органах. Боковые выросты в дальнейшем становятся дорзовентральными. Микрофильные листья имеют лишь один неразветвленный проводящий пучок. При отхождении проводящего пучка от проводящей системы стебля не образуются листовые прорывы (лакуны). Микрофиллы также характерны для псилотообразных.
Сложно устроенные крупные листья (макрофиллы) возникают в результате протекания нескольких взаимосвязанных процессов – агрегации (скучивания), «перевершинивания», уплощения (кладодификации), планации, срастания и редукции некоторых теломов, мезомов или их частей, имевших первоначальное радиальное строение. Агрегация теломов происходила в результате укорачивания, а нередко и редукции мезомов. «Перевершинивание» явилось следствием неравномерного развития стерильных теломов в результате неравной дихотомии. Ось с неограниченным ростом в длину становилась стеблем, а сильно отставшая в росте сдвигалась в сторону и превращалась в боковой орган. Уплощение и срастание происходило в одной или нескольких плоскостях. Под планацией понимают расположение теломов в одной плоскости. Все эти процессы происходили одновременно. Формировались фертильные и стерильные синтеломы, а также синмезомы. Лист как бы захватывает всю надземную часть растения (как у многих современных травянистых папоротников и характеризуется продолжительным верхушечным ростом). Теломные листья являются своего рода «плосковетками». В отличие от энационных, они с самого начала были спороносными, характеризуются сложной системой разветвленных проводящих пучков. При отхождении проводящей системы листа от проводящей системы стебля формируются крупные листовые прорывы, нередко соответствующие веточному прорыву. Макрофилльными являются также листья голосеменных и покрытосеменных. Хвощеобразные по типу формирования листьев занимают промежуточное положение. Их листья возникли из мелких боковых ответвлений надземных осей. Возникновение листьев сильно увеличило поверхность растения. Они принимают на себя функцию фотосинтеза. Это активизировало процессы ассимиляции, газообмена и транспирации.
В ходе эволюции растений из теломов формировались не только листья, но и другие органы высших растений. Например, срастание теломов, расположенных в разных плоскостях, привело к формированию стеблей с более сложным строением стели, чем протостела риниофитов. В процессе эволюции наземных растений наблюдались не только явления усложнения организации, но и явления редукции (уменьшение порядков ветвления, что приводило к упрощению строения органа). Таким путем сформировалась редуцированная листовая пластинка современных хвощей. Регрессивное развитие спорофита в целом наблюдалось у мохообразных. Одним из доказательств является наличие на спорофите некоторых печеночных и антоцеротовых мхов рудиментарных устьиц. При освоении растениями менее влажных местообитаний в процессе адаптации и последующей эволюции размеры листьев уменьшались из-за ослабления их роста. Появляются различные приспособления для ограничения транспирации. Это приводило к расширению адаптационных возможностей растений.
Некоторые листья стали выполнять функцию защиты зоны нарастания стебля и молодых зачатков листьев. Это обусловило появление почек. Сформировался побег (осевая часть – стебель, несущий листья и почки). Дифференциация и усложнение надземных органов, приведшая к формированию побегов, сопровождалась образованием корней в эволюции большинства высших растений для укрепления растения в почве и поглощения воды с растворенными минеральными веществами.
Спорофиллы и трофофиллы (вегетативные листья) согласно теломной теории возникают одновременно и независимо друг от друга. Дальнейшие их развитие шло параллельными путями. Следовательно, независимо друг от друга происходило также формирование вегетативных и спороносных побегов (стробилов). В процессе дальнейшей эволюции высших растений подземные органы совершенствовались, приобретая новые функции, видоизменялись, формируя метаморфозы (корневища, клубни, луковицы и др.). Возникают также новые ароморфозы (семена, цветки, плоды).
Как отмечено, со времени открытия, систематическое положение, классификация, таксономический ранг, объем единиц классификации риниофитов не однократно пересматривались. Это касается большинства групп ископаемых растений благодаря накоплению нового палеонтологического материала.
Некоторые исследователи разделяют риниофиты на 2 или 3 самостоятельных отдела, что не вполне оправдано. Все они связаны рядом важных общих признаков, отличающих их от более организованных высших споровых растений (плаунов, хвощей, папоротников). Они полностью лишены вторичных тканей, корней и побегов; оси всегда протостелические, стела нерасчлененная, спорангии прикрепляются непосредственно к наземным осям (терминально или латерально). Сходство прослеживается и по другим признакам (близость строения спор, ризоидов). Кроме того, существуют так называемые синтетические таксоны (тениокрада – Taeniocrada, куксония, стеганотека – Steganotheca, нотия – Nothia, реналия – Renalia и др.) в которых комбинируются признаки двух или даже трех указанных групп. Они, по-видимому, должны располагаться вместе, в составе одного отдела, в основе филогенетической системы высших растений, подчеркивая их монофилетическое происхождение.
Нередко отдел Риниеобразные условно разделяют на 2 класса – Риниеподобные и Зостеролистоподобные.