Электрические явления в возбудимых тканях
В основе процесса возбуждения лежит изменение потенциала покоя
(ПП) или мембранного потенциала(МП), представляющего собой разность электрических потенциалов внутри и снаружи клетки. МП всегда отрицателен и имеет постоянные значения для каждого типа клеток. Для нейронов потенциал покоя равен минус 70мВ, для мышечного волокна – минус 90 мВ.
Отрицательные значения потенциала покоя связаны с тем, что в покоящейся клетке ионы натрия и калия постоянно перемещаются через клеточную мембрану. При этом К+ из клетки выходит в гораздо большем количестве, чем Na+ входит в клетку, т.к. проницаемость мембраны для ионов К+ примерно в 25 раз больше, чем для ионов Na+. Кроме того, органические анионы из-за своих больших размеров не могут выходить из клетки. В результате в состоянии покоя внутри клетки оказывается больше отрицательных, а снаружи– больше положительных ионов. Возникающая разность потенциалов может быть описана уравнением Нернста:
ПП = 2,3RT/Flg[K+]нap./[K+]внутр.
Из уравнения следует, что, чем выше концентрация калия в среде, тем
меньше величина этого соотношения и меньше соответственно величина МП, т.е. МП имеет калиевую природу. Однако расчетная величина МП все гда ниже, чем есть на самом деле. Например, по расчетам получается, что МП должен быть 90 мВ, а реально фиксируется – 70 мВ. Причина в том, что ионы натрия и хлора тоже вносят свой вклад в создание МП. В состоянии по коя открыта небольшая часть натриевых каналов (20-30 натриевых на 1000 калиевых), соответственно ионы натрия в небольших количествах могут по ступать внутрь клетки. Для поддержания МП на постоянном уровне, т.е. для сохранения ионной асимметрии, служит натрий-калиевый и хлорный насос.
Регистрация МП осуществляется с помощью микроэлектродной техни-
ки (рис. 6). Клетка помещается в камеру, заполненную плазмой крови или физиологическим раствором. Датчиком потенциала является микроэлектрод, представляющий собой стеклянный капилляр с очень тонким кончиком
(диаметром 0,5 мкм), заполненный электролитом (KCl).__
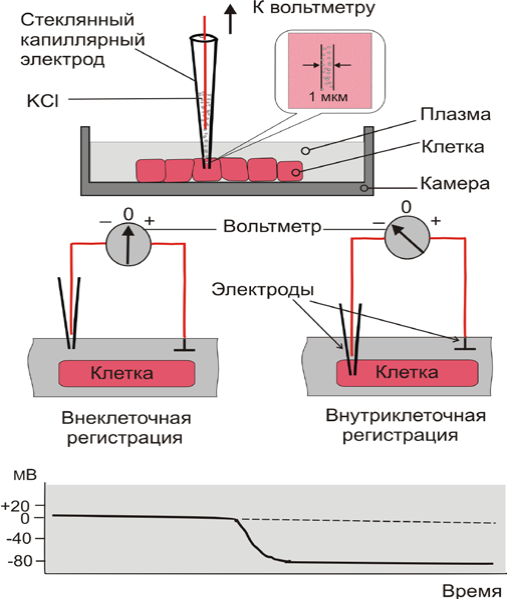
Рис. 6. Техника регистрации мембранного потенциала
Референтным электродом во внеклеточном пространстве служит хло-
рированная серебряная пластинка. Изначально оба электрода находятся во
внеклеточном пространстве, и вольтметр регистрирует между ними нулевую
разность потенциалов. Когда регистрирующий электрод вводят в клетку, т о
вольтметр показывает скачкообразный сдвиг потенциала в отрицательном направлении.
Несмотря на постоянную диффузию ионов МП клеток остается на од-
ном уровне. Следовательно, кроме собственно ионных механизмов формирования мембранного потенциала, связанных с различной проницаемостью клеточной мембраны, имеет место активный механизм его поддержания. Таким механизмом являются ионные насосы, в частности натрий-калиевый насос.
Принцип работы натрий-калиевого насоса.Na/K-насос при каждом
цикле переносит 3Na+ наружу и 2K+ внутрь клетки. Механизм переноса ионов Na+ и K+ через мембрану клетки можно представить следующим образом. Внутри каналоподобной структуры насоса расположены активные цен-тры (места связывания) Na+ и K+, которые поочередно вступают в контакт с внутри- и внеклеточной средой.

Рис. 7. Принцип работы натрий-калиевого насоса: А– связывание Na+ и отщепление K+, т.к. посадочные места в таком положении обладают большим сродством к Na+; Б- связывание молекулы АТФ и фосфорилирование фермента; В– выдвижение посадочных мест во внеклеточную среду; Г- освобождение ионов Na+ и связывание K+ (снижение сродства посадочных мест к Na+ и повышение – к K+); Д– дефосфорилирование АТФазы;Е– возврат системы в первоначальное состояние
Циклическое изменение конформации насоса происходит путем фосфорилирования и дефосфорилирования (присоединение и отщепление остатка фосфорной кислоты) его белковой молекулы (рис. 7). Этот процесс сопровождается изменением сродства мест связывания к соответствующим ионам.
Места связывания, направленные внутрь клетки, имеют низкое сродство к ионам калия и высокое – к ионам натрия. Связывание трех ионов натрия приводит к изменению конформации белка и последующему связыванию АТФ и фосфорилированию фермента. Фосфорилирование фермента приводит к дальнейшему изменению конформации, в результате чего места связывания
оказываются в контакте с внеклеточной средой, теряют сродство к ионам на-
трия и приобретают таковое к ионам калия. Связывание калия вызывает де-
фосфорилирование фермента и возвращает насос в первоначальное положе-
ние, а ионы калия высвобождаются во внутриклеточное пространство.
Потенциал действия.В основе возбуждения нервных и мышечных
клеток лежит повышение проницаемости мембраны для ионов натрия в ре-
зультате открывания натриевых ионных каналов и появления трансмембран-
ных токов, что приводит к быстрому изменению МП в положительном на-
правлении – потенциалу действия (ПД). Таким образом, ПД в отличие от МПимеет натриевую природу. Процесс возбуждения включает в себя генерацию ПД, его распространение и специфический ответ ткани на этот потенциал (сокращение, выделение секрета). Уровень потенциала, при котором деполяризация мембраны приводит к запуску ПД, называется порогом ПДи чаще всего составляет 50 мВ. Разница между МП и порогом ПД называется критическим уровнем деполяризации(КУД). Чем ниже (по абсолютной величине) порог, тем меньше критический уровень деполяризации и выше возбудимость нейрона.
 Рис. 8. Фазы потенциала действия
Рис. 8. Фазы потенциала действия
Потенциал действия(ПД) – это высокоамплитудный, быстро распро-
страняющийся по мембране сигнал, обеспечивающий передачу информации.При регистрации ПД наблюдается типичный пикообразный потенциал, в котором выделяют следующие фазы (рис. 8):
– фазу деполяризации, сопровождающуюся быстрым нарастанием
МП от отрицательных значений до положительного пика – овершута (перелета), составляющего ≈ +30 мВ. То есть это фаза, когда происходит перезарядка мембраны;
– фазу реполяризации, сопровождающуюся восстановлением исход-
ного уровня МП. В ней выделяют фазу быстрой и медленной реполяризации.
Фаза медленной реполяризации представлена следовыми потенциалами –следовой негативностью (гиперполяризацией) и следовой позитивностью(деполяризацией).
Проводимость плазматической мембраны для ионов натрия достигает
максимума гораздо раньше, чем ПД достигает своего пика. Поэтому по достижении пика деполяризации, когда натриевые каналы уже закрыты, а K+ активно диффундирует из клетки, начинается процесс реполяризации. Нарастание проницаемости мембраны для ионов калия происходит гораздо медленнее, чем таковое отмечалось для ионов натрия. Своего максимума она достигает тогда, когда реполяризация уже наполовину завершена. В результате ускоряется вторая фаза реполяризации и возникает следовая гиперполя ризация.
На фоне медленной фазы реполяризации (ее следовой негативности)
происходит активация Na+/K+ насоса и возвращение потенциала мембраны в исходное состояние. Кроме Na+/K+ насоса регуляцию K+ в межклеточном пространстве осуществляют глиальные клетки, обладающие высокой проницаемостью для ионов калия.При действии раздражителя недостаточной силы возникают пассивные
деполяризационные изменения МП – электротонические потенциалы или локальный ответ, амплитуда которого зависит от силы раздражителя (рис. 8).
Рефрактерные периоды
С инактивацией Na+-каналов связана очень важная характеристика возбудимых тканей – рефрактерность – невозбудимость. Закрывшиеся натриевые каналы не сразу восстанавливают свою способность к активации. В связи с этим в течение всей фазы деполяризации потенциала действия и частично фазы реполяризации клетка теряет способность к возбуждению, что именуется фазой абсолютной рефрактерности – состоянием полной невозбудимости(рис. 9).

Рис. 9. Рефрактерные периоды
Постепенно натриевые каналы выходят из состояния инактивации и
возбудимость нервной клетки медленно восстанавливается. Этот период времени носит название относительной рефрактерности. Однако чтобы возбудить клетку в этот период необходимо, увеличить силу раздражения. Во время следовой гиперполяризации, когда потенциал мембраны опускается ниже уровня потенциала покоя и порог возбуждения соответственно увеличивается, имеет место пониженная возбудимость – субнормальность. В случае же следовой деполяризации, когда порог возбудимости понижается, наблюдается повышенная возбудимость – экзальтацияили супернормальность.
Наличие периодов абсолютной рефрактерности ограничивает макси-мальную частоту генерирования ПД. В этой ситуации изменения внешних
сигналов кодируются лишь изменениями частоты ПД (частотный код). Так
как период абсолютной рефрактерности нервной клетки составляет около 1
мс, соответственно, частота их возбуждения (лабильность) может достигать 1000 импульсов в секунду. Однако большинство возбудимых клеток имеет максимальную частоту ПД около 500 импульсов в секунду.
Форма ПД и его длительность зависит от вида возбудимой ткани (рис.
10). ПД нерва составляет около 1 мс, скелетной мышцы – 10 мс, клеток мио-
карда – 200 мс. В одних случаях форма ПД пикообразная, в других – платообразная. Платообразная форма ПД клеток миокарда обеспечивает ритмичность сердечных сокращений.

Рис. 10. Форма и длительность ПД в зависимости от вида возбудимой
ткани