Наследование при предетерминации цитоплазмы
Предетерминацией называется предопределение свойств организмов в последующих поколениях. В ряде случаев наследование признаков связано с особенностями цитоплазмы, возникающими в процессе индивидуального развития организма либо под влиянием факторов внешней среды (онтогенетическая или фенотипическая предетерминация), либо под влиянием генотипа (генотипическая предетерминация).
Онтогенетическая предетерминация. В этом случае наследование некоторых признаков по материнской линии обусловлено изменениями в цитоплазме, возникающими и ней под влиянием определенных внешних факторов. Обычно такие изменения нестойки и через несколько поколений постепенно исчезают, возвращаясь к исходному типу. Например, воздействие повышенной температурой на яйца самок наездника Habrobracon до оплодотворения приводит к изменению окраски тела у их потомства. В последующих поколениях при размножении в нормальных температурных условиях это изменение постепенно затухает. Когда температурному воздействию подвергаются самцы, а самки выращиваются в нормальных условиях, подобного эффекта не наблюдается. Подобные изменения, затухающие в ряду поколений при возвращении организмов в исходные условия, называют длительными модификациями. Механизм их до сих пор не выяснен. Длительные модификации могут постоянно проявляться в ряду поколений при условии сохранения вызывавших их факторов; при отсутствии последних происходит постепенный возврат к исходному состоянию.
Генотипическая предетерминация цитоплазмы происходит под влиянием генотипа материнского организма. Яркий пример – наследование направления завитка раковины у пресноводных гермафродитных моллюсков Limnea. Большинство из них – перекрестно оплодотворяющиеся формы, но некоторые из них способны к самооплодотворению. У этих моллюсков встречаются два типа закручивания раковины: против часовой стрелки (левозакрученные) и по ходу часовой стрелки (правозакрученные). Направление закручивания раковины определяется одной парой аллелей: правозакрученность D доминирует над левозакрученностью d. При реципрокных скрещиваниях гибриды F1, имеющие один и тот же генотип Dd, различаются по фенотипу. В скрещивании ♀ DD × ♂ dd все гибридные особи имеют материнский тип – правозакрученные раковины. В скрещивании ♀ dd × ♂ DD потомство также имеет материнский тип завитка, то есть левозакрученную раковину. От самооплодотворения гетерозиготных форм F1 (Dd) в обоих скрещиваниях все потомки F2 обладают правозакрученной раковиной, хотя гибриды F1 (как и материнские формы) различались по фенотипу. Когда было исследовано потомство от каждой особи F2 в отдельности, то выяснилось, что 1/4 семей имели левый завиток, а 3/4 – правый.
Результаты опытов можно отобразить в виде схемы:
Вариант 1.
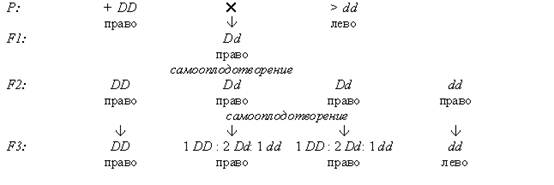
Вариант 2.

Таким образом, простое менделевское расщепление по данной паре признаков 3 : 1 выявилось не в F2, а только в F3. При этом типе наследования фенотип потомков соответствует генотипу матери, а не генотипу зигот, из которых они развиваются. Данный признак предопределяется генотипом материнского организма в цитоплазме яйца в процессе его развития. Рассмотренный тип наследования и является в собственном смысле материнским. Направление завитка раковины определяется характером спирального дробления оплодотворенного яйца, то есть расположением бластомеров по спирали вправо или влево, что, в свою очередь, зависит от ориентации веретена при втором делении дробления.
В данном случае свойства цитоплазмы детерминированы действием хромосомных генов, а не элементами самой цитоплазмы, то есть здесь действует механизм хромосомного наследования, который изменяет цитоплазму яйцеклетки еще до оплодотворения.