Л Е К Ц И Я 10
Тема: Характеристики биоценозов.
(ПРОДУКТИВНОСТЬ и биомасса; ПИРАМИДЫ ЭНЕРГИИ, ЧИСЕЛ И БИОМАСС; СТРУКТУРА; показатели РАЗНООБРАЗИя)
ПРОДУКТИВНОСТЬ и биомасса
Как и для популяций отдельных видов, важным функциональным показателем всего биоценоза, а также его отдельных элементов, является способность к созданию (продуцированию) новых поколений себе подобных, новой биомассы. Это свойство лежит в основе понятия продуктивность.
Первоначальный поток энергии, поступающий на Землю, это солнечное излучение. Наиболее важным для продуктивности биоценозов является суммарная величина потока света напрямую получаемая зелёными растениями. Вообще количество солнечной энергии очень велико и составляет примерно 55 ккал на 1см2 в год. Растения для создания органического вещества используют всего 0,1-0,2% всей солнечной энергии, попадающей на Землю. Остальное затрачивается на нагревание атмосферы, водоёмов, суши и испарение. Но, тем не менее, этого вполне достаточно для поддержания всех жизненных процессов в биосфере Земли. По некоторым данным ежегодно на нашей планете в результате фотосинтеза образуется около 170млрд.т. органического вещества, что в пересчёте на 1га поверхности составляет 3т. Используя солнечную энергию, CO2 и элементы минерального питания, биоценозы продуцируют древесину, листовую массу растений, плоды, животную биомассу и т.п..
Производительность биоценоза, измеряемая количеством органического вещества, которое создано за единицу времени на единице площади, т.е. измеряемая скоростью продуцирования биомассы, называется биологической продуктивностью. Единицы измерения продуктивности – гр/м2 в день, т/км2 в год и т.п.. Продукция любого из трофических уровней – это суммарная продукция всех популяций, занимающих его.
По продуктивности биоценозы разделяют на 5 классов:
Ø Очень высокой биопродуктивности – > 2 кг/м2 в год. Это топические леса; коралловые рифы; морские и океанические побережья, насыщенные жизнью благодаря притоку питательных веществ с континента (речные тростниковые дельты Волги, Дона, Урала).
Ø Высокой биопродуктивности – 1-2 кг/м2 в год. Это липово-дубовые леса, прибрежные заросли рогоза и тростника на озерах, посевы кукурузы и многолетних трав при орошении и удобрении.
Ø Умеренной биопродуктивности – 0,25-1,00 кг/м2 в год. Это многие посевы, сосновые и берёзовые леса, сенокосные луга и степи, заросшие водной растительностью озёра, «морские луга» из водорослей в Японском море.
Ø Низкой биопродуктивности – < 0,25 кг/м2 в год. Это арктические пустыни островов Северного ледовитого океана, тундры, полупустыни Прикаспия, вытоптанные скотом степные пастбища с низким и редким травостоем, горные степи на тонкослойных каменистых почвах. Такая же низкая продуктивность и у большинства океанических и морских глубоководных экосистем.
Ø Очень низкой биопродуктивности – ~ 0 кг/м2 в год. Это те, которые безжизненные, пески настоящих пустынь и льды Антарктики.
Средняя продуктивность биосферы Земли на превышает 0,3 кг/м2 в год., так как на планете преобладают низкопродуктивные экосистемы пустынь и океанов. Общая годовая продукция наземной растительности оценивается ~ 180÷200 млрд.т., а фитомассы океанов ~ 50÷100 млрд.т.. Таким образом, хотя океан занимает ⅔ поверхности Земного Шара, он даёт только ⅓ всей продукции биосферы.
Биопродуктивность биоценоза отличается от запаса биомассы. Некоторые организмы живут много лет (деревья, крупные животные) и их биомасса переходит из года в год как некоторый капитал. Поэтому запас биомассы в лесах ~ в 50 раз, а в степях и на лугах в 3-5 раз больше, чем биопродуктивность. Биомасса леса велика за счёт многолетних частей деревьев – стволов, веток, корней. Поэтому ежегодный прирост (биопродукция) новых листьев, молодых веточек и корней, очередных годичных колец древесины и травяного покрова – составляет не более 2% от этой биомассы (рис.5.1). На лугу запас биомассы значительно меньше – он образован в основном корневищами растений, живущими в почве по нескольку лет. Но прирост зелёной фитомассы за вегетационный период составляет всё же лишь часть от этой биомассы (рис.5.1). В однолетних посевах биопродукция и запас биомассы примерно равны (рис.5.1), так как урожай надземных частей растений, и подземных если это корнеплоды, убирают, а остатки запахивают в почву, где они к весне перегнивают. Как в луговом биоценозе, так и в полевом биоценозе длительность жизни многочисленных почвенных беспозвоночных животных измеряется неделями и месяцами. Их биопродуктивность либо равна запасу биомассы, либо меньше. В водных биоценозах большинство организмов (фотосинтезирующим микроорганизмы, амёбы, инфузории, рачки и др.) живут недолго – всего несколько дней или недель, и за год или даже за лето успевают произвести на свет несколько поколений потомков, т.е несколько своих же биомасс. Поэтому в водоёме в каждый момент времени биомасса организмов меньше, чем их биопродуктивность за вегетационный сезон (рис.5.1). Кроме того, в некоторых водоёмах за счёт того, что рыбы живут по нескольку лет, а жизнь организмов фитопланктона непродолжительна, запас биомассы животных может быть больше запаса биомассы растений. Т.е. если сравнивать биопродуктивность только с фитомассой, то разница будет ещё больше. В морских биоценозах это является правилом (исключая водорослевые луга). При малой фотосинтезирующей массе и недостатке азота и фосфора для фотосинтеза в морях первичная валовая продукция всего в 3 раза ниже, чем в сухопутных биоценозах, хотя биомасса уступает в 350 раз.

Рис. 5.1. Соотношение биомассы и продуктивности различных биоценозов.
Говоря о биопродуктивности следует иметь ввиду следующее:
1. Собственно продуцирование, или синтез нового био вещества, происходит за счёт роста организмов и нарождения новых особей. И тот и другой процессы требуют определённых затрат энергии и вещества.
2. Итоговая чистая продукция всегда меньше общей энергии, полученной организмами с пищей, т.к. некоторая её часть расходуется на дыхание, пищеварение, обогрев тела, т.е на поддержание жизненных функций самого организма, на совершение какой либо работы, и теряется в виде фекалий или при отмирании организмов (потреблённая пища = рост + рождение новых особей + поддержание физиологических функций + работа + фекалии + останки при отмирании).
3. Продуцирование – непрерывный процесс, поэтому при подсчёте общей продукции организмов за тот или иной отрезок времени, необходимо учитывать приросты не только выживших, но и погибших в течении этого периода, особей, ибо они тоже росли и тем самым участвовали в формировании продуктивности сообщества.
4. В сообществе, сохраняющем устойчивое состояние, фактическая продукция данного трофического уровня должна покрывать пищевые потребности организмов следующего уровня. В противном случае общий запас био вещества данного трофического уровня будет неуклонно снижаться из-за выедания. (Как мы убедились ранее, устойчивые адаптированные взаимосвязи между хищниками и жертвами не позволяют первым полностью выедать популяции вторых, за счёт которых они существуют. По это причине эффективность переноса энергии от жертвы к хищнику ослаблена выработанной в процессе эволюции тактикой избегания хищника.)


А) б)
Рис. 5.2. Продуктивность биоценоза.
Вся органическая масса, созданная растениями в единицу времени (обычно за год) называется валовой первичной продукцией (рис.5.2а). Она частично расходуется самими растениями на поддержание жизнедеятельности. Оставшаяся часть – чистая первична продукция (рис.5.2а) – потребляется консументами, или после отмирания перерабатывается редуцентами, а частично осаждается и накапливается в виде запасов торфа, угля, сланцев и т.п.. У большинства растений чистая продукция составляет примерно половину от валовой (рис.5.2б). Первичная продукция, потреблённая консументами частично идёт на создание вторичной продукции (рис.5.2а,б), т.е. прироста их массы в единицу времени. Фитофаги съедают только около 10% биомассы растений. При этом часть потреблённой первичной продукции возвращается в абиотическую среду в виде фекалий, трупов и выделений. И эти вещества могут быть снова потреблены растениями. Поскольку консументы лишь используют ранее созданные органические вещества, вторичную продукцию на валовую и чистую не разделяют. Но её количество также зависит от затрат на жизнедеятельность самих консументов. При интенсивной физической нагрузке (миграции) или при необходимости затрачивать много энергии на согревание тела в холод, продукция (например, рост организмов) уменьшается. Таким образом получается, что при переходе энергии с одного трофического уровня на другой потери составляют примерно 90%. Поэтому вторичная биопродуктивность в 20÷50 раз меньше, чем первичная.
Рассмотрим факторы, определяющие долю чистой первичной продукции, переходящей по «восходящим» трофическим уровням, т.е. эффект переноса вещества и энергии в той или иной трофической цепи :
1. Эффективность потребления – это то, какая часть чистой продукции данного трофического уровня достаётся организмам следующего трофического уровня. В лесах фитофаги потребляют 5% фитопродукции, в степях ~25%, в водных биоценозах, где продуценты – фитопланктон, ~50%. Считается, что хищники из числа позвоночных способны поглощать 50÷100% продукции предыдущего трофического уровня; а если кормом хищникам служат беспозвоночные животные – то лишь 5%.
2. Эффективность усвоения пищи у консументов. Она никогда не бывает равной 1, поскольку часть питательных веществ теряется в виде фекалий. Она низка у фитофагов, детритофагов и потребителей микроорганизмов (20÷50%) и высока у хищников (80%)
3. Эффективность продуцирования – то, какая доля усвоенной пищи и заключённой в ней энергии идёт на создание новой биомассы, потомства. Самая высокая она у микроорганизмов, довольно высока у беспозвоночных животных – 30÷40%, у хладнокровных позвоночных ~10%, у теплокровных она ниже всех – 1÷2%, поскольку большое количество энергии тратится на поддержание постоянной температуры тела.
Если материя (вещества) циркулирует в системе, то энергия нет. Она, использованная хотя бы 1 раз, не может быть снова «пущена в дело». Она может быть только накоплена или трансформирована в другие формы. Экосистема на входе должна иметь постоянный приток энергии.
Законы термодинамики: 1. энергия может переходить из одной формы в другую, но никогда не создаётся вновь и не исчезает. 2. Не может быть ни одного процесса, связанного с превращением энергии, без потери некоторой её части в виде тепла, т.е. нет преобразований энергии со 100% эффективностью.

ПИРАМИДЫ ЭНЕРГИИ, ЧИСЕЛ И БИОМАСС
Итак, по некоторым подсчётам, использование энергии каждым последующим уровнем редко превышает 10%. Поэтому, если представить каждое отдельное звено цепи питания в виде блока, величина которого соответствует продуктивности данного трофического уровня, то получится характерная пирамида – «Пирамида Элтона» (рис.10). Поскольку потери при переходе с уровня на уровень велики, то число звеньев пищевой цепи, а значит этих же самых уровней, ограничено и редко превышает 4. Это называется Правилом Пирамиды чисел. Структура этой пирамиды различна в разных биоценозах. Уменьшение количества доступной энергии обычно влечёт за собой уменьшение численности и биомассы особей на каждом трофическом уровне.

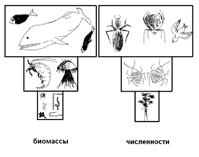

Рис. 5.5. Пирамиды трофических
уровней перевёрнутые
В большинстве наземных биоценозов пирамида биомассы сходна по своему виду с пирамидой энергии – с «повышением» трофического уровня запас биомассы уменьшается. Биомасса растений (наибольшая) превосходит биомассу фитофагов, а биомасса тех, в свою очередь, больше биомассы хищников I порядка, биомасса которых больше биомассы суперхищников (наименьшей). А вот в морских биоценозах, дело обстоит иначе. С повышением трофического уровня запасы биомассы здесь увеличиваются (рис.5.5 биомассы). Ведь, как уже говорилось, основной продуцент здесь – очень мелкий многочисленный фитопланктон, короткоживущий, но с огромной скоростью воспроизводства, т.е. дающий вполне достаточно пищи для последующих трофических уровней. Потребители планктона и далее (по пищевой цепи) хищники живут значительно дольше и накапливают большую собственную биомассу даже при низкой скорости воспроизводства, хотя она и получается меньше общей продуктивности продуцентов. Поэтому пирамида биомассы для таких биоценозов получается как бы перевёрнутой. Впрочем, такая картина может наблюдаться не во все сезоны года. Весной, в период массового развития планктона («цветения воды»), его биомасса может быть выше совокупной биомассы организмов 2-го и 3-го трофических уровней.
Строят ещё пирамиды численности, отражающие плотность особей на каждом трофическом уровне (рис.12.24). Но они менее удобны, поскольку размеры организмов одного уровня сильно различаются. В числе продуцентов в лесу, например, оказываются и дуб и фиалка, а среди фитофагов – и лось и гусеница. Пирамиды численности также могут быть перевёрнутыми в том случае, если продуценты имеют высокую скорость воспроизводства (рис.5.5 численности).
Рассмотренные соотношения блоков пирамид трофических уровней характерны для естественных экосистем, ненарушенных деятельностью человека. В этом случае они сохраняют устойчивость к неблагоприятным внешним воздействиям. Продуктивность экосистемы зависит от ресурсов почвы, атмосферы, солнечного света и тепла. Каждый из этих ресурсов незаменим. Продуктивность в большей мере зависит от того ресурса, которого недостаточно или которого в избытке. Такой ресурс называется лимитирующим фактором. В пустыне урожайность лимитируется количеством осадков, в лесу – содержанием питательных веществ в почве, в тундре – количеством тепла. Чтобы повысить продуктивность экосистем, человек стремится уменьшить влияние лимитирующих факторов – вносит удобрения, орошает, создаёт теплицы и парники. Однако, даже при лучших условиях экосистема не будет продуктивной, или будет давать продукцию за счёт малоценного для человека органического вещества, если её использовать нерационально. Например, изъятие более 30÷50% годового прироста наземной растительности, что может иметь место в сельском и лесном хозяйстве, уменьшает способность экосистемы сопротивляться стрессу. В этой связи следует вспомнить мнение Ю.Одума, который считает, что «Перевыпас скота был одной из причин упадка многих древних цивилизаций.». Хотя понятие «перевыпас» относительно, а «недовыпас» приводит к чрезмерному накоплению детрита и замедлению круговорота минеральных веществ в экосистеме. Поэтому рациональное использование – одно из условий поддержания высокой продуктивности экосистем, вовлечённых человеком в процесс хозяйственной деятельности. Биопродуктивность может снижаться и при загрязнении окружающей среды отходами и токсикантами.
СТРУКТУРА
Видовая. Это состав видов и их обилие. Чем больше видов, значит больше экологических ниш, т.е. выше богатство среды. Богатые видами биоценозы приурочены к тем местам, которые находятся в оптимальных условиях абиотической среды – тропические леса, коралловые рифы, долины рек. Видовая структура зависит также от истории формирования современной флоры и фауны. Поэтому молодые, недавно сформировавшиеся биоценозы имеют более бедный видовой состав, чем давно сложившиеся, зрелые. Так же обеднены по сравнению со зрелыми природными биоценозы, созданные человеком (поля, сады, огороды). Основные экологические процессы определяются, конечно, наиболее многочисленными видами.
Пространственная. Определяется главным образом пространственным распределением растительной части биоценоза. Выделяют вертикальное (ярусность) и горизонтальное (мозаичность) сложение фитоценоза. Оно является результатом конкуренции и позволяет наиболее полно использовать ресурсы среды, прежде всего свет, тепло и влагу. В лесах умеренных широт чётко выделяют следующие ярусы – древесный (первый и второй); подлесок из кустарников; травянистый и моховый, подстилка. Мозаичность образуется в результате неоднородности микрорельефа, почвы, средообразующей деятельности растений и животных. Сочетание различных растительных ассоциаций создаёт многообразие мест для обитания животных. Животное население биоценоза, привязанное к растительности, также распределено по вертикальным ярусам и горизонтальным районам, областям. Наиболее богата видами лесная подстилка. Достаточно чётко приурочены к ярусам определённые группы насекомых, птицы при строительстве гнезда и поиске корма.
Экологическая. Это соотношение различных экологических групп организмов по тому или иному признаку, входящих в состав биоценоза. Биоценозы, имеющие сходную экологическую структуру, могут, тем не менее, сильно различаться по видовому составу. Экологические ниши могут быть заняты даже неродственными видами, но имеющими одинаковые биоценотические функции.
Биоморфологический спектр. Это состав и соотношение слагающих сообщество жизненных форм. Определяется типом сообщества (луг, лес, море, горные тундры, пустыни и т.п.). Число жизненных форм, входящих в биоценоз, обычно гораздо больше числа образующих его видов. Но набор жизненных форм, занимающих разные экологические нишы, всегда более закономерен для данного местообитания и гораздо лучше характеризует экологические условия в сообществе как абиотические, так и биотические.
Итак. Биоморфологический спектр и пространственная структура сообщества являются показателями имеющегося в данном местообитании разнообразия экологических ниш, богатства и полноты использования сообществом ресурсов среды; а также показателем относительной устойчивости сообщества и степени антропогенного влияния на него.
Состав трофической сети. Отражает число трофических уровней, соотношение продуцентов и консументов, первичных, вторичных, третичных.
ПОКАЗАТЕЛИ РАЗНООБРАЗИЯ
Виды, которые наиболее обильно представленные в биоценозе, занимающие ведущее, господствующее положение здесь, называются доминантами. Но не все доминантные виды одинаково влияют на биоценоз. Здесь, как правило, есть и так называемые эдификаторы – виды, которые своей жизнедеятельностью в основном и создают среду для всего сообщества и без которых в связи с этим существование большинства других видов невозможно. Это строители сообщества, в подавляющем большинстве случаев, конечно же, растения. Удаление вида-эдификатора из биоценоза влечёт за собой изменение даже физической среды в первую очередь микроклимата биотопа. Ярким примером эдификаторов являются лесообразующие породы: есть леса сосновые, берёзовые, дубовые, елово-пихтовые, эвкалиптовые и т.п. В некоторых случаях эдификаторами являются и животные. На территориях занятыми «мощными» колониями сурков, например, именно их роющая деятельность определяет большей частью характер ландшафта, микроклимат и условия произрастания растений.
Как известно, любой биоценоз состоит из множества организмов, каждый из которых занимает свой определённое функциональное место в общем круговороте вещества и энергии, в общей системе взаимосвязей внутри биоценоза. Разнообразие организмов или их групп называется биоразнообразием. Оно определяется богатством видов этих организмов или их групп (списочный состав) и их количественным соотношением. Чем выше это богатство и чем равномернее по обилию (количественно) представлены различные виды или группы видов, тем выше разнообразие. А если численно будет преобладать один вид, а все остальные будут малочисленны или редки, то разнообразие будет низким. Различают видовое разнообразие, разнообразие экологических групп, а также разнообразие самих экосистем. Биоразнообразие в конкретном смысле – это количественный показатель, связывающий число группировок живых систем, выделенных по тому или иному параметру и критерию, и выровненность их обилия по этим группировкам.
Выражают разнообразие или в виде графика, где по оси абсцисс откладывают, например, виды в порядке убывания их численности, а по оси ординат численность либо класс, ранг обилия этих видов и в дальнейшем по протяжённости, форме и крутизне получившейся кривой судят об уровне разнообразия; или в виде математических индексов, когда по определённым формулам рассчитывают соотношение между числом группировок живых систем, выделенных по различным параметрам и критериям, и показателями их доли в общем обилии. Можно инвентаризировать число и соотношение элементов определенной живой системы в пределах местообитания, района, ландшафта или большой группы территорий (a-, g-, e-разнообразие соответственно, табл.5.1); а можно изучать изменение видового состава, структуры и разнообразия живых систем вдоль градиента среды или градиента местообитаний, сравнивая между собой территории a-, g-разнообразия, области e-разнообразия, оценивая степень их сходства по уровню разнообразия (b-, D-, Ω-разнообразие соответственно) (это будет дифференцирующее разнообразие, табл.5.1); то есть изучать разнообразие живых систем в различном масштабе. В настоящее время для некоторых категорий, в основном для a и b, разработано значительное количество алгебраических (индексов разнообразия, индексы сходства) и графических показателей (модели распределения группировок живых систем по обилию). К сожалению, к единому общеупотребляемому выражению разнообразия учёные ещё не пришли.
Таблица 5.1
Формы и типы разнообразия по Р.Уиттекеру [1960, 1977] и др.
| Инвентаризационное разнообразие | Дифференцирующее разнообразие |
| Альфа-разнообразие (разнообразие внутри местообитания) | Бета-разнообразие (разнообразие между местообитаниями) |
| Гамма-разнообразие (для ландшафта или геогр.области, зоны) | Дельта-разнообразие (между географическими районами) |
| Эпсилон-разнообразие (для биома, географического региона, включающего различные ландшафты) | Омега-разнообразие (между биомами, географическими регионами, континентами) |